
慢性炎症管理(专业版)
炎症是机体的防御机制,构成人体免疫反应的一部分。如果没有炎症反应,感染、伤口和组织损伤等都将无法自愈。然而,炎症因子持续存在并且损伤组织却是发生慢性炎症的根本原因。
英文名称:Chronic Inflammation Management
慢性炎症是人类“无声的”杀手,因缺乏临床症状至今仍没有医疗措施。慢性炎症涉及到绝大部分的慢性疾病。因此,慢性炎症管理成为欧美对慢性病管理的主要方法之一。本文介绍了炎症产生原因和风险因素,慢性炎症的干预路径和综合干预方案。
1.急性炎症:这是对组织损伤或感染的适应性免疫反应。
2.慢性炎症(Chronic Inflammation):由细胞应激和及其功能失调引起。这是低度炎症,可通过多种机制导致疾病和与年龄有关的退化。
1). 维生素D:
维生素D似乎通过抑制促炎性前列腺素和抑制炎症介质NF-κB发挥抗炎活性1。尽管缺乏对其在人类中抗炎活性的干预研究,但几项观察性研究表明,维生素D缺乏可能会促进炎症。维生素D缺乏症在炎症性疾病(包括类风湿性关节炎、炎症性肠病、系统性红斑狼疮和糖尿病)患者中比在健康人中更常见2。在易发生低水平炎症的人群中也更常见3,手术后维生素D水平可能下降(一种与急性炎症相关的情况),伴随着CRP的升高4。
在一项对548名心力衰竭患者的研究中,维生素D水平低与CRP升高有关5,在一组46名患有内皮功能障碍的中年男性中,与IL-6和NF-κB的升高有关6。
2). 维生素E:
维生素E在体内起抗氧化剂的作用。具体来说,维生素E被掺入低密度脂蛋白(LDL)颗粒中,并保护它们免受氧化损伤;它似乎也通过其他机制预防动脉粥样硬化7。维生素E的γ-生育酚形式似乎补充了α-生育酚的抗炎作用。γ-生育酚已被证明可抑制COX-2并减弱IL-1β信号传导8,9。
在一项针对代谢综合征受试者的小型临床试验中,与安慰剂相比,γ-生育醇和α-生育酚的组合有效抑制了C反应蛋白和TNF-α水平10。在本研究中,两种生育酚的组合比单独使用任何一种都要好,这促使研究人员指出,“在氧化应激和炎症的生物标志物方面,α-生育酚和γ-生育酚的联合补充似乎优于单独补充,需要在前瞻性临床试验中进行测试……”。
3). 镁:
在两项大型观察研究(妇女健康倡议和哈佛护士研究)中,镁摄入量越高,hs-CRP、IL-6和TNF-α受体(衡量TNF-α活性的指标)越低11,12。来自动脉粥样硬化多民族研究的数据未能发现镁摄入量最高和最低的个体之间IL-6或CRP水平的显著差异,但确实发现饮食中较高的镁与较低水平的炎症相关蛋白同型半胱氨酸和纤维蛋白原之间存在显著关联13。
在饮食炎症指数中,镁被评为最抗炎的饮食因子,该指数根据人类和动物的实验和观察数据,对42种常见的饮食成分进行了评级,以降低CRP水平14。
4). 锌和硒:
含锌和硒的抗氧化蛋白(如超氧化物歧化酶和谷胱甘肽过氧化物酶)可减少活性氧(自由基),从而间接抑制NF-κB活性,并阻止几种炎症酶和细胞因子的产生。锌还能以更直接的方式抑制NF-κB15,16。在儿童和老年人等容易缺锌的人群中,补充锌与炎症的减少有关17,18。在几项研究中,适量补充锌可以降低老年人的低水平炎症和循环促炎因子(CRP、TNF-α、IL-6和IL-8)得到了降低19-21。与锌一样,硒缺乏在与疾病相关的慢性炎症状态(如败血症)中很常见22,144补充硒与减少炎症和改善患者预后有关16。
5).鱼油:
鱼油是ω-3脂肪酸二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)的最佳来源,这些脂肪酸只能在人类中有限程度地合成。Omega-3脂肪酸在预防数万名患者的心血管疾病和死亡率方面已经得到了很好的研究;ω-3的抗炎作用有助于这种活性23。在许多其他炎症性疾病的研究中,特别是哮喘、炎症性肠病和类风湿性关节炎,ω-3也被证明能成功地改善患者的预后24,25。
几项大型观察性试验的数据证实了鱼油/ω-3摄入量增加与全身炎症减少之间的联系。在来自卫生专业人员随访研究的855名健康参与者中,ω-3脂肪酸的摄入与血浆TNF-α活性标志物水平较低有关;有趣的是,高摄入ω-3和ω-6脂肪酸(通常被认为是促炎性的)与炎症水平最低有关26。由727名女性组成的护士健康研究I队列显示,在摄入ω-3最多的20%人群中,炎症标志物(包括CRP和IL-6)的浓度较低27。在ATTICA对3000多名没有任何心血管疾病证据的希腊男性和女性进行的研究中,每周食用300g以上鱼类的参与者平均CRP降低33%,IL-6降低33%,并且TNF-α比不吃鱼的参与者低21%28。在来自动脉粥样硬化多民族研究(MESA)队列的5677名没有心血管疾病的男性和女性样本中,长链ω-3的摄入(来自鱼类或补充剂)与多种炎症标志物(包括CRP、IL-6和TNF-α受体,一种衡量TNF-α活性的指标)的血浆浓度降低有关29。
6). 特异性促分解介质(SPMs):
特异性促分解介质(SPMs)是来源于多不饱和脂肪酸代谢的细胞信号分子。它们在解决炎症反应中起着重要作用30,31。SPMs包括脂氧素(lipoxins)、消炎素(Resolvins)、保护素(Protectins)和马雷素(Maresins)及其前体物。它们不仅抑制炎症,而且积极解决炎症过程,帮助组织恢复正常状态31。SPMs招募巨噬细胞清除细胞碎片,从而恢复健康细胞32;使炎症损伤的组织恢复活力33;并恢复抗炎细胞因子和促炎细胞因子之间的平衡34,35。
慢性炎症会导致衰老和变性,SPMs可能有助于降低与炎症和衰老相关的某些疾病的风险31,36-39。对肥胖小鼠的研究表明,低度炎症会导致代谢紊乱,SPMs前体可以减轻肥胖并发症38。另一项研究表明,在喂食高胆固醇饮食九周的小鼠中,每天1-5mg/kg的SPMs Resolvin E1可以减少动脉粥样硬化病变37。
源自深海鱼油的SPMs有助于支持身体有效解决炎症反应。身体从ω-3脂肪酸中产生SPMs,包括EPA和DHA40;然而,它们并没有完全转化为SPMs。Omega-3和SPMs在体内也有不同的作用。
为了更好地了解补充鱼油(EPA和DHA)对SPMs血浆脂质代谢物浓度的影响,进行了一项新的临床试验。共有21名患有慢性炎症的患者被随机分组,依次接受3g/天的EPA或DHA,为期10周;其间有10周的洗脱期。这项研究还包括一项离体分析,评估了EPA和DHA衍生的SPMs对炎症细胞(单核细胞)的影响有何不同41。
结果表明,EPA和DHA都以各自的方式提高了血浆中SPMs和SPM前体的水平。在研究的离体部分,从参与者的血液样本中提取的单核细胞(巨噬细胞前体细胞)在体外用毒素(LPS)刺激以诱导炎症反应。EPA和DHA在体外均能降低促炎因子TNF-α和单核细胞趋化蛋白-1(MCP-1)的基因表达。EPA对促炎IL-6的表达没有影响;然而,DHA降低了它。DHA降低了抗炎IL-10的表达;然而,EPA却增加了它。有趣的是,DHA比EPA更有效地抑制单个的促炎细胞因子,但EPA更有效地平衡炎症细胞因子与抗炎细胞因子IL-10的比例。
这些结果为EPA和DHA的独立作用提供了新的见解,并表明这两种ω-3脂肪酸在炎症缓解中发挥着重要作用。
2. 抑制炎症的功能性营养:
1).脱氢表雄酮(DHEA):
性激素水平低与炎症标记物的系统性增加有关42。作为一种肾上腺类固醇激素,DHEA是性类固醇睾酮和雌激素的前体。DHEA在年轻人中含量丰富,但随着年龄的增长而稳步下降,可能是与年龄相关的性类固醇减少的部分原因43。在细胞培养和动物模型中,DHEA可以抑制炎症细胞因子活性,在某些情况下比睾酮或雌激素更有效44。慢性炎症本身可能会降低DHEA水平45。在一项小型研究中,老年志愿者补充DHEA(50mg/天,持续两年)可显著降低TNF-α和IL-6水平,降低内脏脂肪量,改善糖耐量(二者均与炎症有关)46。
2).姜黄素:
广泛的体外和动物研究已经检验了姜黄素对实验诱导的炎症性疾病(动脉粥样硬化、关节炎、糖尿病、肝病、胃肠道疾病和癌症)和疾病标志物(脂氧合酶、环氧合酶、TNF-α、IL-1β、NF-κB等)的影响47,48。研究姜黄素对炎症性疾病患者预后影响的人类研究较少,但大多数姜黄素的小型随机对照试验始终显示,患者在几种炎症性疾病方面有所改善,包括银屑病、肠易激综合征(IBS)、类风湿性关节炎和炎症性眼病49,50。
3).白藜芦醇和紫檀芪:
白藜芦醇发挥抗炎活性的确切机制尚未确定,尽管它在动物模型和人类细胞培养中抑制多种促炎化合物(环加氧酶、TNF-α、IL-1β、IL-6、NF-κB)51,52。其相关化合物紫檀芪在细胞培养中显示出类似的炎症标志物抑制作用53。
炎症免疫反应的调节可能有助于白藜芦醇在心脏病、癌症、急性胰腺炎和炎症性肠病动物模型中的保护作用54。白藜芦醇也可以预防一般的、低水平的副炎症:在对10名健康志愿者进行的一项小型交叉研究中,当与一顿高脂肪、高碳水化合物(930千卡)一起服用时,白藜芦醇(100mg)可以防止餐后氧化和炎症标志物的急剧增加。例如,在试验餐后5小时内,IL-1β的合成增加了91%;使用白藜芦醇时,这种增加明显较少(29%)55。
4).茶多酚:
数十项体外和动物研究证实了绿茶和红茶多酚的抗炎作用56。绿茶多酚EGCG和茶黄素通过抑制NF-κB信号通路发挥抗炎作用,它降低了细胞培养实验中几种炎症蛋白(脂氧合酶、环氧合酶、TNF-α、IL-1β、IL-6和IL-8)的表达57。EGCG还抑制组胺的产生和释放,组胺是过敏和炎症反应的关键介质58。
在喝茶的观察性研究中,与不喝茶的人相比,每天喝2杯茶(红茶或绿茶)可使CRP降低近20%,并显著降低其他两种炎症标志物(血清淀粉样蛋白a和半抗原,在冠心病中升高)的水平59。在临床干预中,红茶在降低炎症标志物方面似乎比绿茶更成功11。117在一项小型试验中,健康的非吸烟者服用红茶提取物(相当于每天四杯茶)六周,CRP也降低了25%60。在一项大型研究中,对患有冠心病的高风险健康人也观察到了类似的平均降低,但在CRP起始值最高(>3 mg/L)的个体中,CRP降低了40-50%61。
5). 乳香:
乳香是阿育吠陀医学中一种传统的抗关节炎药物;其抗炎特性被归因于对5-LOX的特异性抑制和乳香酸(乳香主要活性成分)减少促炎白三烯的产生62。在细胞培养中,粗提物和高度纯化的乳香提取物都抑制了促炎TNF-α和IL-1β的产生63。其中一种乳香酸乙酰-11-酮- β -乳香酸(AKBA)在小鼠中是NF-κB活性的抑制剂64;而在啮齿动物炎症模型中,四种最丰富的乳香酸的局部混合物可以减轻炎症65。
最近对乳香治疗炎症条件的人体试验的系统综述表明,对该提取物的少量随机对照试验为其用于哮喘和骨关节炎产生了令人鼓舞的结果66。因此需要进行更大规模的研究来证实提取物是一种有效的治疗方法。标准化乳香提取物(30%AKBA)在减轻骨关节炎患者的疼痛方面是有效的67。当与非挥发性乳香精油结合使用时,标准化提取物(AprèsFlex或Aflapin)在较低浓度下表现出更高的活性68。尽管结果不一,但乳香提取物用于炎症性肠病的用途已在多项临床试验中进行了研究69-71。
6). 类胡萝卜素:
在妇女健康和衰老研究中,血液中α-胡萝卜素和总类胡萝卜素水平最高的参与者在研究开始时比类胡萝卜素水平较低的参与者有更低的IL-6水平72。血液中α-和β-胡萝卜素、叶黄素/玉米黄质或总类胡萝卜素水平最低的参与者在2年的时间里更有可能经历IL-6的增加。
7). N-乙酰半胱氨酸(NAC):
NF-κB通路的激活在炎症细胞因子基因的激活中起着核心作用;NAC在细胞培养中抑制NF-κB,降低IL-6和IL-8等细胞因子的表达73,74。确定NAC在降低人类慢性炎症方面的作用的数据有限,但显示出前景。在慢性肾脏病患者中,补充NAC八周后,循环IL-6水平略有下降,但具有统计学意义75。研究开始时,对有严重炎症的人的影响更为明显(通过hs-CRP测量)。在一项针对烧伤患者的小型研究中,NAC还降低了全身炎症标志物76。
8). 菠萝蛋白酶:
菠萝蛋白酶是来源于菠萝茎的包括蛋白水解酶的混合物。在临床前研究中,菠萝蛋白酶一直表现出多种抗炎特性77,78。这些抗炎作用归因于COX-2活性降低和前列腺素合成减少。菠萝蛋白酶还通过对纤维蛋白和纤维蛋白原的作用调节凝血78。
菠萝蛋白酶治疗炎症条件的人体试验也产生了有希望的结果79。一项对四项研究的荟萃分析发现,菠萝蛋白酶在牙科手术后可以减轻肿胀,并具有镇痛和抗炎作用80。
一项为期一个月的开放标签研究评估了不同剂量菠萝蛋白酶对轻度急性膝关节疼痛的影响。77名膝盖疼痛不到三个月但身体健康的参与者每天服用200或400mg菠萝蛋白酶。参与者在基线和一个月后通过标准化问卷评估自己的疼痛和功能。与基线相比,200mg组的症状评分改善了41%,400mg组改善了59%。两组患者的心理健康状况也显著改善81。
一项随机对照非劣效性试验在103名膝骨关节炎患者中比较了含有菠萝蛋白酶(90mg,每日三次)加胰蛋白酶和芸香苷的标准化酶制剂与非甾体抗炎药双氯芬酸(Voltaren等)(50mg,每日二次)。研究人员发现,菠萝蛋白酶在患者报告的疼痛和功能评估中的作用至少与双氯芬酸一样好80。
另一项随机试验招募了30名有症状的颞下颌关节炎患者,比较了三种治疗方案:
第1组和第3组的反应被认为是相同的,再次证明含有菠萝蛋白酶的组合与双氯芬酸一样有效。然而,与单独使用双氯芬酸或口服酶治疗的参与者相比,第2组的参与者在疼痛和炎症方面有显著改善82。这些发现表明菠萝蛋白酶和这些其他成分可以补充非甾体抗炎药在炎症条件的管理。
一项针对49名骨关节炎患者的试点研究发现,食用含有乳香和菠萝蛋白酶混合物的补充剂长达六个月,可以显著提高自我报告的生活质量分数。据报道,基线时受影响最严重的关节症状改善最为明显83。
在另一项随机对照非劣效性试验中,研究人员将90名患有疼痛性髋关节骨关节炎的患者分为两组:一半接受含有菠萝蛋白酶、胰蛋白酶和芸香苷的口服酶制剂治疗六周,另一半接受抗炎药双氯芬酸治疗。他们发现,菠萝蛋白酶制剂在疼痛、僵硬和身体功能的标准化评分中与双氯芬酸一样有效,而且往往比药物对照品耐受性更好。此外,研究人员报告称,71%服用含菠萝蛋白酶制剂的受试者和61%服用双氯芬酸的受试人员的疗效为“良好”或“非常好”77,78。
菠萝蛋白酶与歌手等声带炎症:
菠萝蛋白酶等纯天然的抗炎药可能对歌手经常出现的声带炎症有用。
歌手、教师和其他经常使用或过度使用声音的人(有时被称为误用甚至滥用)有很高的声带紧张风险,这可能会导致炎症和肿胀84-86。这种慢性刺激会导致息肉和结节的发展87,88。这些问题通常可以通过休息声音(如果可能的话)、正确使用声音、口腔补水和使用室内加湿器来解决;在某些情况下,可能需要口服或注射皮质类固醇,甚至手术切除84-86,88,89。当导致病变发展的活动(如唱歌或大喊大叫)持续时,这些病变复发并不罕见。因此,经常寻求非侵入性的自然支持。
根据支持菠萝蛋白酶减轻炎症能力的证据,菠萝蛋白酶作为治疗声带肿胀和炎症的药物在歌手和声乐家中广受欢迎90。考虑到非甾体抗炎药是导致声带问题常见原因的风险因素,这一点尤为重要和有价值88,91。
一项临床研究对56名因良性声带疾病接受手术的患者进行了检查,发现术后使用菠萝蛋白酶(80mg)和乳香(750mg)联合治疗,每天两次,持续一周,可有效促进组织修复和嗓音恢复。与吸入类固醇治疗相比,这种治疗具有良好的耐受性,产生的副作用更少87。
9). 芝麻木脂素:
观察到芝麻油可以减少真菌和大鼠肝细胞中花生四烯酸的产生,从而鉴定出芝麻木脂素(芝麻脂素/Sesamin、芝麻林素/Sesamolin、芝麻明酚/Sesaminol)是合成花生四烯酸所使用的酶之一Δ5去饱和酶(δ -5去饱和酶)的特异性抑制剂92。通过抑制Δ5去饱和酶,芝麻木脂素可以减少促炎前列腺素、白三烯和凝血素的合成,这些都需要花生四烯酸作为起始物质93。
在动物模型中,高芝麻油的饮食减少了促炎前列腺素PGE-1和-2以及血栓素B2的产生94。在人类中,五周的芝麻素补充(39mg/天)减少了促炎血管收缩剂20-羟基二十碳四烯酸(20-HETE,酶5-LOX的产物)减少30%95。芝麻木脂素的这种潜在抗炎特性可能部分解释了其观察到的降压活性96。
10).黑籽油:
黑籽油(或称黑孜然籽油)是一种抗氧化剂和抗炎剂,传统上用于促进消化、皮肤和肝脏健康。261研究表明,黑籽油平衡免疫和炎症反应,支持健康的免疫系统功能97,98。随着年龄的增长,破坏性炎症途径增加、免疫反应受损,这与关节炎、过敏和心血管疾病等慢性疾病有关99。
黑籽油及其活性成分百里香醌(Thymoquinone)被认为可以抑制有害炎症,同时支持抵抗感染和癌症所需的正常免疫反应98,100。黑籽油可刺激巨噬细胞和辅助T细胞的活性101。
在一项安慰剂对照试验中,40名患有类风湿性关节炎的女性每天服用安慰剂一个月,然后每天两次服用500mg黑籽油、持续一个月。据报道,关节炎症状,包括关节肿胀和晨僵,治疗后疾病严重程度有所下降102。在一项前瞻性双盲研究中,66名过敏性鼻炎患者暴露于黑孜然籽油中30天。结果鼻炎的症状,包括充血、鼻子发痒和打喷嚏,在治疗后有所减轻103。动物研究表明,补充黑籽油具有心脏保护作用,因为它可以减少脂质氧化,促进健康的胆固醇水平104。
3. 支持线粒体:
线粒体呼吸过程中产生的活性氧会导致炎症,如上述有关内容所述。衰老个体特别容易受到线粒体相关氧化应激的影响,因为线粒体随着年龄的增长而变得越来越功能障碍。采取措施支持线粒体的完整性和效率可以帮助减轻线粒体功能不良引起的一些系统氧化和炎症负担。辅酶Q10和吡咯喹啉醌(PQQ)这两种营养素是强大的线粒体保护剂,研究支持这些化合物的抗炎作用105,106。
1).吡咯喹啉醌(PQQ):
PQQ是对细胞能量稳态和氧化还原平衡至关重要的酶的辅因子107。多项研究表明,PQQ在线粒体环境应激和氧化负荷增加期间发挥保护作用106,108。在一项研究中,给予补充PQQ的饮食的大鼠表现出更大的能量消耗,肝组织线粒体密度增加。补充PQQ的大鼠甘油三酯也较低,与未服用PQQ的鼠相比,它们的心脏更能防止缺氧109。在心脏组织氧气供应有限的时期,氧化应激和随后的炎症损伤细胞急剧上升;该动物模型的研究表明,PQQ可以通过在不利条件下保持线粒体效率来避免这种炎症细胞破坏。
2). 辅酶Q10:
CoQ10是线粒体ATP产生过程中不可或缺的中间产物。研究表明,在炎症条件下,辅酶Q10水平较低。在一项研究中,感染性休克患者的辅酶Q10水平明显低于健康人,并且在患者中,较低的CoQ10水平与较高水平的一种炎症介质VCAM相关110。
在动物模型中,给大鼠饮用添加果糖的饮用水,该实验会导致肥胖、糖尿病和其他炎症并发症。补充辅酶Q10可通过降低肝脏中CRP和其他炎症介质的表达来减轻炎症反应111。实验室研究表明,辅酶Q10调节数百个基因的表达,其中许多与炎症信号有关112。特别重要的是,一项实验表明,在生理相关浓度下,CoQ10能够通过调节NF-κB信号通路,将诱导的TNF-α减弱25%以上112。
4. 预防炎症性糖化反应:
已知升高的血糖和糖基化终产物(AGEs)在引发炎症风暴中的作用。幸运的是,除了减少热量摄入以抑制空腹和餐后葡萄糖浓度外,一些天然化合物还能改善糖基化过程,并可能有助于控制糖诱导的炎症级联反应。在这些抗糖基化营养素中,最主要的是B族维生素家族成员苯磷硫胺和一种氨基酸肌肽。
1). 苯磷硫胺:
自20世纪90年代中期以来,苯磷硫胺一直被用于治疗糖尿病并发症113。最近的证据继续支持它作为一种强大的保护剂来对抗血糖诱导的组织损伤。在一项临床试验中,165名糖尿病受试者被随机分配接受每天300或600mg苯磷硫胺或安慰剂治疗六周。干预期结束后,服用苯磷硫胺的患者表现出剂量依赖性的神经性疼痛改善114。一个动物模型发现,苯磷硫胺通过有效抑制炎症来缓解神经性疼痛115。此外,实验室研究表明,除了阻断糖基化反应外,苯磷硫胺可能通过调节COX和LOX酶活性更直接地调节炎症116。
2). 肌肽:
肌肽在体内发挥一系列有利的生物化学作用。它能有力地减缓糖基化反应并缓解氧化应激117。此外,几项实验表明,肌肽具有显著的抑制各种细胞类型炎症的能力118-120。不幸的是,肌肽水平在10岁至70岁之间下降了63%121。此外,在2型糖尿病患者中,骨骼肌肌肽含量明显低于健康对照受试者122。当肌肽作为化学诱导糖尿病动物的补充剂时,它能够保护脆弱的视网膜细胞免受与高血糖相关的炎症并发症的影响123。
3). 绞股蓝:
绞股蓝在亚洲医学中用于治疗多种健康状况,包括血脂异常、2型糖尿病和炎症124。它的作用至少部分是由于它能够激活一种单磷酸腺苷活化蛋白激酶(AMPK)的关键酶。这种影响葡萄糖代谢和脂肪储存的酶被称为“代谢主开关”,因为它控制着从食物中提取能量并在全身储存和分配能量的许多途径125。
超重对AMPK激活和慢性炎症有显著影响;它抑制AMPK的激活,导致腹部脂肪沉积,进而激活全身炎症。同时,炎症本身抑制AMPK的激活,产生粘性循环126。
然而,更多的AMPK激活有助于减肥并可以抑制炎症127-129。此外,AMPK激活增加与肝脏脂肪积聚减少有关,这是炎症化学物质的另一个来源130。
有证据表明绞股蓝具有抗炎作用。在一项实验室研究中,研究人员发现该草药提取物能显著抑制几种炎症化学物质,包括TNF-α、白细胞介素-6和COX-2信使核糖核酸(mRNA)131。另一项针对24名2型糖尿病患者的研究发现,饮用该草药泡茶12周可显著降低胰岛素抵抗,而胰岛素抵抗是导致全身炎症的关键因素132,133。
4).橙皮苷:
橙皮苷和相关黄酮类化合物存在于多种植物中,尤其是柑橘类水果的果皮中134,135。橙皮苷的消化会产生一种称为陈皮素(Hesperetin)的化合物以及其他代谢产物。这些化合物是强大的自由基清除剂,具有抗炎、胰岛素增敏和降脂活性136,137。动物和体外研究结果表明,橙皮苷对血糖和脂质水平的积极作用可能部分与AMPK途径的激活有关138-140。越来越多的证据表明,橙皮苷可能有助于预防和治疗与衰老相关的许多慢性疾病136。
橙皮苷可以部分通过激活AMPK信号通路来预防糖尿病及其并发症。巧合的是,二甲双胍,一种领先的糖尿病药物,也激活AMPK通路。在一项针对24名糖尿病参与者的为期六周的随机对照试验中,每天补充500mg橙皮苷可以改善血糖控制,提高总抗氧化能力,减少氧化应激和DNA损伤141。另一组研究人员使用尿陈皮素作为膳食橙皮苷的标志物,发现摄入橙皮苷水平最高的人在4.6年内患糖尿病的风险比摄入橙皮苷水平最低的人低32%142。
在一项随机对照试验中,24名患有代谢综合征的成年人每天服用500mg橙皮苷或安慰剂治疗三周。洗脱期后,用橙皮苷重复试验,安慰剂分配颠倒。橙皮苷治疗改善了内皮功能,这可能是其有益于心血管系统的重要机制之一。相对于安慰剂,补充橙皮苷还导致炎症标志物hs-CRP的中位水平降低33%,总胆固醇、载脂蛋白B(apoB)和血管炎症标志物的水平显著降低139。在另一项针对有证据表明存在血管功能障碍的超重成年人的随机对照试验中,连续六周每天补充450mg橙皮苷可降低血压,降低血管炎症标志物143。另一项对照临床试验包括75名心脏病发作患者,他们被随机分配接受每天600mg橙皮苷或安慰剂治疗四周。服用橙皮苷的患者的高密度脂蛋白胆固醇、血管炎症标志物、脂肪酸和葡萄糖代谢水平显著改善144。
据于上述慢性炎症干预路径及相关营养素或功能营养,其综合干预方案如下:
以及参阅本网站如下专文了解更多内容:
参考文献:
1. Krishnan AV et al. The role of vitamin D in cancer prevention and treatment. Endocrinol Metab Clin North Am. 2010;39(2):401–18
2. Guillot X et al. Vitamin D and inflammation. Joint Bone Spine. 2010;77(6):552–557
3. Awad AB et al. Vitamin D and metabolic syndrome risk factors: evidence and mechanisms. Crit Rev Food Sci Nutr. 2012;52(2):103–112
4. Reid D et al. The relation between acute changes in the systemic inflammatory response and plasma 25-hydroxyvitamin D concentrations after elective knee arthroplasty. American Journal of Clinical Nutrition. 2011;93(5):1006–1011
5. Liu LCY et al. Vitamin D status and outcomes in heart failure patients. Eur. J. Heart Fail. 2011;13(6):619–625
6. Jablonski KL et al. 25-Hydroxyvitamin D deficiency is associated with inflammation-linked vascular endothelial dysfunction in middle-aged and older adults. Hypertension. 2011;57(1):63–69
7. Meydani M. Vitamin E and atherosclerosis: beyond prevention of LDL oxidation. J Nutr. 2001Feb;131(2):366S-8S.
8. Jiang Q et al. gamma-tocopherol and its major metabolite, in contrast to alpha-tocopherol, inhibit cyclooxygenase activity in macrophages and epithelial cells. Proc Natl Acad Sci USA. 2000 Oct 10;97(21):11494-9.
9. Sjoholm A et al. gamma-tocopherol partially protects insulin-secreting cells against functional inhibition by nitric oxide. Biochem Biophys Res Commun. 2000 Oct 22;277(2):334-40.
10. Devaraj S et al. Gamma-tocopherol supplementation alone and in combination with alpha-tocopherol alters biomarkers of oxidative stress and inflammation in subjects with metabolic syndrome. Free Radic Biol Med. 2008;44(6):1203-8.
11. Galland L. Diet and inflammation. Nutr Clin Pract. 2010;25(6):634–640
12. Chacko S et al. Relations of dietary magnesium intake to biomarkers of inflammation and endothelial dysfunction in an ethnically diverse cohort of postmenopausal women. Diabetes. 2010;
13. de Oliveira Otto et al. Dietary micronutrient intakes are associated with markers of inflammation but not with markers of subclinical atherosclerosis. Journal of Nutrition. 2011;141(8):1508–1515
14. Cavicchia PP et al. A new dietary inflammatory index predicts interval changes in serum high-sensitivity C-reactive protein. Journal of Nutrition. 2009;139(12):2365–2372
15. Prasad AS. Zinc: role in immunity, oxidative stress and chronic inflammation. Current Opinion in Clinical Nutrition and Metabolic Care. 2009;12(6):646–652
16. Duntas LH. Selenium and inflammation: underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009;41(6):443–447
17. Kelishadi R et al. Effect of zinc supplementation on markers of insulin resistance, oxidative stress, and inflammation among prepubescent children with metabolic syndrome. Metab Syndr Relat Disord. 2010;8(6):505–510
18. Wong CP et al. Zinc and its role in age-related inflammation and immune dysfunction. Mol. Nutr. Food Res. 2011;
19. Bao B et al. Zinc decreases C-reactive protein, lipid peroxidation, and inflammatory cytokines in elderly subjects: a potential implication of zinc as an atheroprotective agent. American Journal of Clinical Nutrition. 2010;91(6):1634–1641
20. Kahmann L et al. Zinc supplementation in the elderly reduces spontaneous inflammatory cytokine release and restores T cell functions. Rejuvenation Res. 2008;11(1):227–237
21. Mariani E et al. Simultaneous evaluation of circulating chemokine and cytokine profiles in elderly subjects by multiplex technology: relationship with zinc status. Biogerontology. 2006;7(5-6):449–459
22. Maehira F et al. Alterations of serum selenium concentrations in the acutephase of pathological conditions. Clin. Chim. Acta. 2002;316(1-2):137–146
23. Marik PE et al. Omega-3 dietary supplements and the risk of cardiovascular events: a systematic review. Clin Cardiol. 2009;32(7):365–372
24. Calder PC. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am J Clin Nutr. 2006;83(6 Suppl):1505S–1519S
25. Giugliano D et al. The Effects of Diet on Inflammation. J Am Coll Cardiol. 2006;48(4):677–685
26. Pischon T et al. Habitual dietary intake of n-3 and n-6 fatty acids in relation to inflammatory markers among US men and women. Circulation. 2003;108(2):155–160
27. Lopez-Garcia E et al. Consumption of (n-3) fatty acids is related to plasma biomarkers of inflammation and endothelial activation in women. J Nutr. 2004;134(7):1806–1811
28. Zampelas A et al. Fish consumption among healthy adults is associated with decreased levels of inflammatory markers related to cardiovascular disease: the ATTICA study. J Am Coll Cardiol. 2005;46(1):120–124.
29. He K et al. Associations of dietary long-chain n-3 polyunsaturated fatty acids and fish with biomarkers of inflammation and endothelial activation (from the Multi-Ethnic Study of Atherosclerosis [MESA]). Am J Cardiol. 2009;103(9):1238–1243
30. Serhan CN et. Novel anti-inflammatory--pro-resolving mediators and their receptors. Current topics in medicinal chemistry. 2011;11(6):629-647.
31. Basil MC et al. Specialized pro-resolving mediators: endogenous regulators of infection and inflammation. Nature Reviews Immunology. 2016;16(1):51-67.
32. Serhan CN. Treating inflammation and infection in the 21st century: new hints from decoding resolution mediators and mechanisms. Faseb j. 2017;31(4):1273-1288.
33. Hasturk H et al. Resolvin E1 regulates inflammation at the cellular and tissue level and restores tissue homeostasis in vivo. J Immunol. 2007;179(10):7021-7029.
34. Serhan CN. Pro-resolving lipid mediators are leads for resolution physiology. Nature. 2014;510(7503):92-101.
35. Serhan CN. Discovery of specialized pro-resolving mediators marks the dawn of resolution physiology and pharmacology. Mol Aspects Med. 2017;58:1-11.
36. Wang X et al. Insufficient resolution response in the hippocampus of a senescence-accelerated mouse model--SAMP8. J Mol Neurosci. 2015;55(2):396-405.
37. Salic K et al. Resolvin E1 attenuates atherosclerosis in absence of cholesterol-lowering effects and on top of atorvastatin. Atherosclerosis. 2016;250:158-165.
38. Neuhofer A et al. Impaired local production of proresolving lipid mediators in obesity and 17-HDHA as a potential treatment for obesity-associated inflammation. Diabetes. 2013;62(6):1945-1956.
39. Huang J et al. Targeting the D Series Resolvin Receptor System for the Treatment of Osteoarthritis Pain. Arthritis Rheumatol. 2017;69(5):996-1008.
40. Norris PC et al. Identification of specialized pro-resolving mediator clusters from healthy adults after intravenous low-dose endotoxin and omega-3 supplementation: a methodological validation. Scientific Reports. 2018;8(1):18050.
41. So J et al. EPA and DHA differentially modulate monocyte inflammatory response in subjects with chronic inflammation in part via plasma specialized pro-resolving lipid mediators: A randomized, double-blind, crossover study. Atherosclerosis. 2020;316:90-98.
42. Singh T et al. Inflammatory markers in population studies of aging. Ageing Res Rev. 2011;10(3):319–329
43. Heffner KL. Neuroendocrine effects of stress on immunity in the elderly: implications for inflammatory disease. Immunol Allergy Clin North Am. 2011;31(1):95–108
44. Gordon CM et al. Adrenal and gonadal steroids inhibit IL-6 secretion by human marrow cells. Cytokine. 2001;16(5):178–186
45. Ernestam et al. Increased DHEAS levels in patients with rheumatoid arthritis after treatment with tumor necrosis factor antagonists: evidence for improved adrenal function. The Journal of Rheumatology. 2007;34(7):1451–1458
46. Weiss EP et al. Dehydroepiandrosterone (DHEA) replacement decreases insulin resistance and lowers inflammatory cytokines in aging humans. Aging. 2011;3(5):533–542
47. Chainani-Wu N. Safety and anti-inflammatory activity of curcumin: a component of turmeric (Curcuma longa). J Altern Complement Med. 2003;9(1):161–168
48. Bengmark S. Curcumin, an atoxic antioxidant and natural NFkappaB, cyclooxygenase-2, lipooxygenase, and inducible nitric oxide synthase inhibitor: a shield against acute and chronic diseases. JPEN J Parenter Enteral Nutr. 2006;30(1):45–51
49. Epstein J et al. Curcumin suppresses p38 mitogen-activated protein kinase activation, reduces IL-1beta and matrix metalloproteinase-3 and enhances IL-10 in the mucosa of children and adults with inflammatory bowel disease. Br J Nutr. 2010;103(6):824–832
50. White B et al. Clinical Inquiry. Does turmeric relieve inflammatory conditions? J Fam Pract. 2011;60(3):155–156
51. Jha RK et al. Emerging role of resveratrol in the treatment of severe acute pancreatitis. Front Biosci (Schol Ed). 2010;2:168–175
52. Khanduja KL et al. Resveratrol inhibits N-nitrosodiethylamine-induced ornithine decarboxylase and cyclooxygenase in mice. J. Nutr. Sci. Vitaminol. 2004;50(1):61–65
53. Pan Z et al. Identification of molecular pathways affected by pterostilbene, a natural dimethylether analog of resveratrol. BMC Med Genomics. 2008;1:7
54. Clarke JQ et al. A review of complementary and alternative approaches to immunomodulation. Nutrition in Clinical Practice. 2008;23(1):49–62
55. Ghanim H et al. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J Clin Endocrinol Metab. 2011;96(5):1409–1414
56. Singh R et al. Green tea polyphenol epigallocatechin-3-gallate: inflammation and arthritis. [corrected]. Life Sci. 2010;86(25-26):907–918
57. de Mejia et al. Bioactive components of tea: cancer, inflammation and behavior. Brain Behav. Immun. 2009;23(6):721–731
58. Melgarejo E et al. Targeting of histamine producing cells by EGCG: a green dart against inflammation? J. Physiol. Biochem. 2010;66(3):265–270
59. De Bacquer D et al. Epidemiological evidence for an association between habitual tea consumption and markers of chronic inflammation. Atherosclerosis. 2006;189(2):428–435
60. Steptoe A et al. The effects of chronic tea intake on platelet activation and inflammation: a double-blind placebo controlled trial. Atherosclerosis. 2007;193(2):277–282
61. Bahorun T et al. Black tea reduces uric acid and C-reactive protein levels in humans susceptible to cardiovascular diseases. Toxicology. 2010;278(1):68–74
62. Boswellia serrata. Altern Med Rev. 2008;13(2):165–167
63. Gayathri B et al. Pure compound from Boswellia serrata extract exhibits anti-inflammatory property in human PBMCs and mouse macrophages through inhibition of TNFalpha, IL-1beta, NO and MAP kinases. International Immunopharmacology. 2007;7(4):473–482
64. Cuaz-Pérolin C et al. Antiinflammatory and antiatherogenic effects of the NF-kappaB inhibitor acetyl-11-keto-beta-boswellic acid in LPS-challenged ApoE-/- mice. Arteriosclerosis, Thrombosis, and Vascular Biology. 2008;28(2):272–277
65. Singh S et al. Boswellic acids: A leukotriene inhibitor also effective through topical application in inflammatory disorders. Phytomedicine. 2008;15(6-7):400–407
66. Ernst E. Frankincense: systematic review. BMJ. 2008;337:a2813
67. Sengupta K et al. A double blind, randomized, placebo controlled study of the efficacy and safety of 5-Loxin for treatment of osteoarthritis of the knee. Arthritis Res. Ther. 2008;10(4):R85
68. Sengupta K et al. Comparative efficacy and tolerability of 5-Loxin and AflapinAgainst osteoarthritis of the knee: a double blind, randomized, placebo controlled clinical study. Int J Med Sci. 2010;7(6):366–377
69. Gupta I et al. Effects of Boswellia serrata gum resin in patients with ulcerative colitis. Eur. J. Med. Res. 1997;2(1):37–43
70. Gupta I et al. Effects of gum resin of Boswellia serrata in patients with chronic colitis. Planta Med. 2001;67(5):391–395
71. Holtmeier W et al. Randomized, placebo-controlled, double-blind trial of Boswellia serrata in maintaining remission of Crohn's disease: good safety profile but lack of efficacy. Inflamm. Bowel Dis. 2011;17(2):573–582
72. Walston J et al. Serum antioxidants, inflammation, and total mortality in older women. Am. J. Epidemiol. 2006;163(1):18–26
73. Araki et al. N-acetylcysteine inhibits induction of nitric oxide synthase in 3T3-L1 adipocytes. J. UOEH. 2007;29(4):417–429
74. Radomska-Leśniewska et al. Influence of N-acetylcysteine on ICAM-1 expression and IL-8 release from endothelial and epithelial cells. J. Physiol. Pharmacol. 2006;57 Suppl 4:325–334
75. Nascimento MM et al. Effect of oral N-acetylcysteine treatment on plasma inflammatory and oxidative stress markers in peritoneal dialysis patients: a placebo-controlled study. Perit Dial Int. 2010;30(3):336–342
76. Csontos C et al. Effect of N-acetylcysteine treatment on the expression of leukocyte surface markers after burn injury. Burns. 2011;37(3):453–464
77. de Lencastre Novaes LC et al. Stability, purification, and applications of bromelain: A review. Biotechnol Prog. Jan-Feb 2016;32(1):5-13.
78. Chakraborty AJ et al. Bromelain a Potential Bioactive Compound: A Comprehensive Overview from a Pharmacological Perspective. Life (Basel). Apr 6 2021;11(4)
79. Colletti A et al. Recent Advances and Insights into Bromelain Processing, Pharmacokinetics and Therapeutic Uses. Applied Sciences. 2021;11(18):8428.
80. de Souza GM et al. Is bromelain effective in controlling the inflammatory parameters of pain, edema, and trismus after lower third molar surgery? A systematic review and meta-analysis. Phytother Res. Mar 2019;33(3):473-481.
81. Walker AF et al. Bromelain reduces mild acute knee pain and improves well-being in a dose-dependent fashion in an open study of otherwise healthy adults. Phytomedicine. Dec 2002;9(8):681-6.
82. Jayachandran S et al. Efficacy of Bromelain along with Trypsin, Rutoside Trihydrate Enzymes and Diclofenac Sodium Combination Therapy for the treatment of TMJ Osteoarthritis - A Randomised Clinical Trial. Journal of clinical and diagnostic research : JCDR. Jun 2017;11(6):ZC09-ZC11.
83. Italiano G et al. Benefits of a Food Supplement Containing Boswellia serrata and Bromelain for Improving the Quality of Life in Patients with Osteoarthritis: A Pilot Study. Journal of alternative and complementary medicine (New York, NY). Feb 2020;26(2):123-129.
84. National Institute on Deafness and Other Communication Disorders. Taking Care of Your Voice. Updated 4/15/2021. https://www.nidcd.nih.gov/health/taking-care-your-voice
85. Centre ICE. The voice and its disorders in teachers. INSERM Collective Expert Reports. Institut national de la santé et de la recherche médicale. Copyright © 2000-, Institut national de la santé et de la recherche médicale (INSERM).
86. Baylor University. How to Prevent and Repair Vocal Damage. Updated 3/26/2020, https://onlinegrad.baylor.edu/resources/how-to-prevent-and-repair-vocal-damage-for-teachers-fitness-instructors-podcast-hosts-more/
87. Zambito P et al. THE EFFICACY OF EVAN® IN POST-OPERATIVE TREATMENT OF BENIGN CORDAL INJURIES: AN OBSERVATIONAL PILOT STUDY. Accessed 09/28/2021, http://www.frontieraorl.it/en/772-2/
88. Hancock RL et al. Vocal fold hemorrhage: when vocal fold blood vessels are not up to the task! Voice and Speech Review. 2014/01/02 2014;8(1):36-39.
89. Al-Ali M et al. The role of steroid injection for vocal folds lesions in professional voice users. J Otolaryngol Head Neck Surg. Jul 20 2020;49(1):50.
90. Insuan O et al. Anti-Inflammatory Effect of Pineapple Rhizome Bromelain through Downregulation of the NF-kappaB- and MAPKs-Signaling Pathways in Lipopolysaccharide (LPS)-Stimulated RAW264.7 Cells. Current issues in molecular biology. May 7 2021;43(1):93-106.
91. Paknezhad H et al. Vocal Fold Hemorrhage. Ear, Nose & Throat Journal. 2021/07/01 2019;100(6):NP276-NP277.
92. Shimizu S et al. Sesamin is a potent and specific inhibitor of delta 5 desaturase in polyunsaturated fatty acid biosynthesis. Lipids. 1991;26(7):512–516
93. Harikumar KB et al. Sesamin manifests chemopreventive effects through the suppression of NF-kappa B-regulated cell survival, proliferation, invasion, and angiogenic gene products. Mol. Cancer Res. 2010;8(5):751–761
94. Chavali SR et al. Decreased production of interleukin-1-beta, prostaglandin-E2 and thromboxane-B2, and elevated levels of interleukin-6 and -10 are associated with increased survival during endotoxic shock in mice consuming diets enriched with sesame seed oil supplemented with Quil-A saponin. Int. Arch. Allergy Immunol. 1997;114(2):153–160
95. Wu et al. Inhibition of 20-hydroxyeicosatetraenoic acid synthesis using specific plant lignans: in vitro and human studies. Hypertension. 2009;54(5):1151–1158
96. Miyawaki T et al. Antihypertensive effects of sesamin in humans. J. Nutr. Sci. Vitaminol. 2009;55(1):87–91
97. Srinivasan K. Cumin (Cuminum cyminum) and black cumin (Nigella sativa) seeds: traditional uses, chemical constituents, and nutraceutical effects. Food Quality and Safety. 2018;2(1):1-16.
98. Padhye S et al. From here to eternity - the secret of Pharaohs: Therapeutic potential of black cumin seeds and beyond. Cancer Ther. 2008;6(b):495-510.
99. Chung HY et al. Molecular inflammation: underpinnings of aging and age-related diseases. Ageing Res Rev. 2009;8(1):18-30.
100. Salem ML. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int Immunopharmacol. 2005;5(13-14):1749-1770.
101. Salem ML et al. Protective effect of black seed oil from Nigella sativa against murine cytomegalovirus infection. Int J Immunopharmacol. 2000;22(9):729-740.
102. Gheita TA et al. Effectiveness of Nigella sativa oil in the management of rheumatoid arthritis patients: a placebo controlled study. Phytotherapy research: PTR. 2012;26(8):1246-1248.
103. Nikakhlagh S et al. Herbal treatment of allergic rhinitis: the use of Nigella sativa. American journal of otolaryngology. 2011;32(5):402-407.
104. Al-Naqeep G et al. Antiatherogenic Potential of Nigella sativa Seeds and Oil in Diet-Induced Hypercholesterolemia in Rabbits. Evidence-based complementary and alternative medicine: eCAM. 2011;2011:213628.
105. Sourris KC et al. Ubiquinone (coenzyme Q10) prevents renal mitochondrial dysfunction in an experimental model of type 2 diabetes. Free Radic Biol Med. 2012 Feb 1;52(3):716-23.
106. Tao R et al. Pyrroloquinoline quinone preserves mitochondrial function and prevents oxidative injury in adult rat cardiac myocytes. Biochem Biophys Res Commun. 2007 Nov 16;363(2):257-62.
107. Rucker R et al. Potential physiological importance of pyrroloquinoline quinone. Altern Med Rev. 2009 Sep;14(3):268-77.
108. Xiong XH et al. Production and radioprotective effects of pyrroloquinoline quinone. Int J Mol Sci. 2011;12(12):8913-23.
109. Bauerly K et al. Altering pyrroloquinoline quinone nutritional status modulates mitochondrial, lipid, and energy metabolism in rats. PLoS One. 2011;6(7):e21779.
110. Donnino MW et al. Coenzyme Q10 levels are low and may be associated with the inflammatory cascade in septic shock. Crit Care. 2011 Aug 9;15(4):R189.
111. Sohet FM et al. Coenzyme Q10 supplementation lowers hepatic oxidative stress and inflammation associated with diet-induced obesity in mice. Biochem Pharmacol. 2009 Dec 1;78(11):1391-400.
112. Schmelzer C et al. Functions of coenzyme Q10 in inflammation and gene expression. Biofactors. 2008;32(1-4):179-83.
113. Stracke H et al. A benfotiamine-vitamin B combination in treatment of diabetic polyneuropathy. Exp Clin Endocrinol Diabetes. 1996;104(4):311-6.
114. Stracke H et al. Benfotiamine in diabetic polyneuropathy (BENDIP): results of a randomised, double blind, placebo-controlled clinical study. Exp Clin Endocrinol Diabetes. 2008 Nov;116(10):600-5.
115. Sanchez-Ramirez GM et al. Benfotiamine relieves inflammatory and neuropathic pain in rats. Eur J Pharmacol. 2006 Jan 13;530(1-2):48-53.
116. Shoeb M et al. Anti-inflammatory effects of benfotiamine are mediated through the regulation of the arachidonic acid pathway in macrophages. Free Radic Biol Med. 2012 Jan 1;52(1):182-90.
117. Vistoli G et al. Transforming dietary peptides in promising lead compounds: the case of bioavailable carnosine analogs. Amino Acids. 2012 Jul;43(1):111-26.
118. Fleisher-Berkovich S et al. Inhibitory effect of carnosine and N-acetyl carnosine on LPS-induced microglial oxidative stress and inflammation. Peptides. 2009 Jul;30(7):1306-12.
119. Tsai SJ. Antioxidative and Anti-Inflammatory Protection from Carnosine in the Striatum of MPTP-Treated Mice. J Agric Food Chem. 2010 Oct 6.
120. Boldyrev AA et al. [Carnosine: endogenous physiological corrector of antioxidative system activity]. Usp Fiziol Nauk. 2007 Jul-Sep;38(3):57-71.
121. Hipkiss AR. On the enigma of carnosine’s anti-ageing actions. Exp Gerontol. 2009 Apr;44(4):237-42.
122. Gualano B et al. Reduced muscle carnosine content in type 2, but not in type 1 diabetic patients. Amino Acids. 2011 Nov 27.
123. Pfister F et al. Oral carnosine supplementation prevents vascular damage in experimental diabetic retinopathy. Cell Physiol Biochem. 2011;28(1):125-36.
124. Gauhar R et al. Heat-processed Gynostemma pentaphyllum extract improves obesity in ob/ob mice by activating AMP-activated protein kinase. Biotechnology letters. 2012;34(9):1607-1616.
125. Winder WW et al. AMP-activated protein kinase, a metabolic master switch: possible roles in type 2 diabetes. Am J Physiol. 1999 Jul;277(1 Pt 1):E1-10.
126. Ruderman NB et al. AMPK, insulin resistance, and the metabolic syndrome. J Clin Invest. 2013 Jul 1;123(7):2764-72.
127. Park SH et al. Antiobesity effect of Gynostemma pentaphyllum extract (actiponin): a randomized, double-blind, placebo-controlled trial. Obesity (Silver Spring). 2014 Jan;22(1):63-71.
128. Towler MC, Hardie DG. AMP-activated protein kinase in metabolic control and insulin signaling. Circ Res. 2007 Feb 16;100(3):328-41.
129. Salminen A et al. AMP-activated protein kinase inhibits NF-kappaB signaling and inflammation: impact on healthspan and life span. J Mol Med (Berl). 2011 Jul;89(7):667-76.
130. Bijland S et al. Role of AMP-activated protein kinase in adipose tissue metabolism and inflammation. Clin Sci (Lond). 2013 Apr;124(8):491-507.
131. Xie Z et al. Chemical composition and anti-proliferative and anti-inflammatory effects of the leaf and whole-plant samples of diploid and tetraploid Gynostemma pentaphyllum (Thunb.) Makino. Food chemistry. 2012;132(1):125-133.
132. Bastard J-P et al. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur. Cytokine Netw. 2006;17(1):4–12
133. Huyen VT et al. Antidiabetic effect of Gynostemma pentaphyllum tea in randomly assigned type 2 diabetic patients. Horm Metab Res. 2010 May;42(5):353-7.
134. Umeno A et al. Antioxidative and Antidiabetic Effects of Natural Polyphenols and Isoflavones. Molecules. May 30 2016;21(6).
135. Devi KP et al. Hesperidin: A promising anticancer agent from nature. Industrial Crops and Products. 2015;76:582-589.
136. Li C et al. Health-promoting effects of the citrus flavanone hesperidin. Critical reviews in food science and nutrition. Feb 11 2017;57(3):613-631.
137. Roohbakhsh A et al. Neuropharmacological properties and pharmacokinetics of the citrus flavonoids hesperidin and hesperetin--a mini-review. Life sciences. Sep 15 2014;113(1-2):1-6.
138. Jia S et al. Hypoglycemic and hypolipidemic effects of neohesperidin derived from Citrus aurantium L. in diabetic KK-A(y) mice. Food Funct. Mar 2015;6(3):878-886.
139. Rizza S et al. Citrus polyphenol hesperidin stimulates production of nitric oxide in endothelial cells while improving endothelial function and reducing inflammatory markers in patients with metabolic syndrome. The Journal of clinical endocrinology and metabolism. May 2011;96(5):E782-792.
140. Zhang J et al. Purification of naringin and neohesperidin from Huyou (Citrus changshanensis) fruit and their effects on glucose consumption in human HepG2 cells. Food chemistry. Dec 01 2012;135(3):1471-1478.
141. Homayouni F, Haidari F, Hedayati M, Zakerkish M, Ahmadi K. Hesperidin Supplementation Alleviates Oxidative DNA Damage and Lipid Peroxidation in Type 2 Diabetes: A Randomized Double-Blind Placebo-Controlled Clinical Trial. Phytotherapy research. Aug 14 2017.
142. Sun Q et al. Urinary Excretion of Select Dietary Polyphenol Metabolites Is Associated with a Lower Risk of Type 2 Diabetes in Proximate but Not Remote Follow-Up in a Prospective Investigation in 2 Cohorts of US Women. The Journal of nutrition. Jun 2015;145(6):1280-1288.
143. Salden BN et al. Randomized clinical trial on the efficacy of hesperidin 2S on validated cardiovascular biomarkers in healthy overweight individuals. The American journal of clinical nutrition. Dec 2016;104(6):1523-1533.
144. Haidari F et al. Hesperidin supplementation modulates inflammatory responses following myocardial infarction. Journal of the American College of Nutrition. 2015;34(3):205-211.
美国国立卫生研究院
http://www.nih.gov
美国国立公众健康网
www.medlineplus.gov
英国今日医学新闻网
www.medicalnewstoday.com
免责声明和安全信息
概述
炎症是机体的防御机制,构成人体免疫反应的一部分。如果没有炎症反应,感染、伤口和组织损伤等都将无法自愈。然而,炎症因子持续存在并且损伤组织却是发生慢性炎症的根本原因。慢性炎症是年轻机体功能丧失和退行性疾病发展的主要因素,包括自体免疫病、心血管病、糖尿病和老年痴呆症等。慢性炎症是人类“无声的”杀手,因缺乏临床症状至今仍没有医疗措施。慢性炎症涉及到绝大部分的慢性疾病。因此,慢性炎症管理成为欧美对慢性病管理的主要方法之一。本文介绍了炎症产生原因和风险因素,慢性炎症的干预路径和综合干预方案。
炎症产生原因
一般认为,炎症是组织损伤的急性或短期反应,其产生特征性的症状并通常能自发愈合。然而,慢性炎症则将导致多种慢性疾病,包括心脑血管病、糖尿病、慢阻肺病和老年痴呆,以及某些癌症和类风湿关节炎等。1.急性炎症:这是对组织损伤或感染的适应性免疫反应。
- 急性炎症反应由下列因素引发:
- 组织损伤,例如创伤、烫伤或化学物灼伤等。
- 病毒、细菌、真菌或寄生虫感染等
- 急性炎症的表现特征:
- 发红、发热:流向创伤部位的血流量增加所致
- 肿胀:由受伤部位积聚的液体引起
- 疼痛:由肿胀压缩受伤部位的神经末梢而导致疼痛。疼痛使得生物体意识到组织受伤,是一种保护机制。
- 急性炎症的保护作用包括:
- 防止传染源扩散和对附近组织的损害
- 有助于清除受损的组织和病原体
- 协助身体的修复过程
急性炎症反复发作,而发作间期无明显症状也是慢性炎症,如慢性胆囊炎、慢性肾盂肾炎等。
2.慢性炎症(Chronic Inflammation):由细胞应激和及其功能失调引起。这是低度炎症,可通过多种机制导致疾病和与年龄有关的退化。
- 线粒体功能障碍:线粒体是机体“能源站”,人体活力好取决于最佳状态的线粒体功能。然而,线粒体产能过程也导致自由基不断累积,引起线粒体损伤和功能失调,进而诱发破坏细胞和组织的炎性介质,导致细胞凋亡。如果失控,则可能导致癌症。
- 外源性因素如环境毒素、烟草和压力等及内源性因素如活性氧都可引起细胞应激。
- 衰老及线粒体效率下降与自由基产生增加有关。
- 循环糖:主要是血液中的葡萄糖和果糖。它们可与蛋白质和脂质接触,产生糖基化终产物(AGEs),结合细胞膜受体(RAGE)后可激活炎症介质核因子(NF-kB),后者移向细胞核激活众多炎症基因。除高血糖外,高温烹饪的食物尤其是红肉中的AGEs也可激活炎症基因。
- 其他诱导慢性炎症的物质:
炎症标志物和介质
应激诱发的炎症一旦被触发,促炎细胞因子及其慢性作用(低度炎症)可能持续存在数年、数十年而引起细胞死亡,并与多种慢性疾病形成有关。- 肿瘤坏死因子α(TNF-α):可通过多种类型的免疫细胞(如巨噬细胞)释放,以响应细胞损伤、应激或感染,是生理性炎症反应、免疫监视和抗肿瘤等所必需的。然而,过量的TNF-α可导致慢性炎症状态,增加血栓形成并降低心脏收缩力,与肿瘤的发生和发展可能有关。
- 核因子κB(NF-kB):在炎症反应启动中很重要。当细胞暴露于损伤信号(如TNF-α或氧化应激)时,它们可激活NF-κB,从而开启涉及炎症反应的400多个基因的表达。其中包括其他炎症细胞因子和促炎酶,例如环氧合酶2(COX-2)和脂氧合酶(LOX)。COX-2是负责合成促炎性前列腺素的酶,也是非甾体类抗炎药NSAIDs(布洛芬、阿司匹林等)和COX-2抑制剂(西乐葆)的标靶。
- 白细胞介素/白介素:主要由白细胞产生的在促进和解决炎症中具有许多功能的细胞因子。白介素在信息传递、激活与调节免疫细胞,介导T、B细胞活化、增殖与分化及在炎症反应中起重要作用。包括IL-1β,IL-6和IL-8:
- IL-1β帮助免疫细胞移出血管,进入受损或功能失调的组织。
- IL-6具有促炎和抗炎作用,并协调急性炎症的进展和消退过程中所需化合物的产生。
- IL-8由免疫细胞和非免疫细胞表达,并有助于将中性粒细胞(破坏病原体的免疫细胞)吸引到损伤部位。
- C-反应蛋白(CRP):一种在炎症急性反应期间由肝脏快速产生的几种蛋白质之一,主要是覆盖受损的细胞,使其更易于被其他免疫细胞识别。高水平的CRP与心血管疾病、卒中等风险密切关系。因此,是体检的一种常用指标。
- 类花生酸类物质(Eicosanoids):包括前列腺素、前列环素、白三烯和血栓素等,由靠近炎症部位的细胞产生形成免疫防疫。
- 前列腺素除了在炎症反应中作用外,在细胞生长、肾功能、消化以及血管收缩和扩张中具有多种功能。
- 白三烯:在炎症反应期间招募和激活白细胞具有重要促炎作用,并在气道收缩和过敏反应中作用。
- 血栓素是凝血过程的重要介体。
类花生酸类物质由细胞膜中的不饱和脂肪酸合成,主要原料是亚油酸ω-6及其衍生物花生四烯酸(AA)等。omega-3脂肪酸可平衡亚油酸作用,与降低炎症和心血管疾病风险的生物标志物有关
- 环氧合酶(COX)和脂氧合酶(LOX):这是类花生酸合成所必需的酶。
- COX促成ω-6衍生物转化为许多前列腺素或血栓素之一。阿司匹林和非甾体抗炎药(布洛芬、萘普生)的镇痛和抗炎活性是由于其对COX酶的抑制,从而降低炎症、发烧和疼痛。
- LOX将脂肪酸转化为白三烯,后者是重要的炎症介质,可放大炎症效应。因此,LOX是抗炎治疗的重要靶标之一。
虽然COX和LOX通常与促炎过程相关,但两种酶也产生抑制并促进组织修复(包括前列环素和脂氧素)的因子。COX和LOX酶从促炎活性到消炎活性的正确过渡对于健康炎症反应的进展很重要。
产生炎症的风险因素
可能促进慢性炎症的危险因素包括如下:- 年龄增长,老年体内促炎细胞因子如IL-6和TNF-α较高,与氧化损伤累积、线粒体功能障碍、性激素水平下降和内脏脂肪多等有关。
- 肥胖:脂肪组织是一种内分泌器官,可储存和分泌多种激素和细胞因子进入循环系统,如TNF-α和IL-6,并影响全身的新陈代谢。
- 内脏脂肪细胞可产生大量的炎性因子。
- 巨噬细胞浸润脂肪组织与BMI指数成正比,并产生促炎因子,引起胰岛素抵抗和代谢综合征。
- 高脂肪、高热量饮食:与更高的促炎症标志物相关,尤其在肥胖和糖尿病人群中。反式脂肪引起的炎症标志物在超重人群中增加可能更为显著。热量限制提供强大的抗炎保护,是缓解生理压力的有效手段。热量限制法是一种有效的抗衰老方法,更多可参阅本网专文:热量限制法与抗衰老 >>
- 性激素水平低:性激素参与调节免疫/炎症反应。中性粒细胞和巨噬细胞等免疫细胞具有雌激素和雄激素的受体,能够响应性激素变化水平。睾丸激素和雌激素可抑制细胞因子(IL-1β,IL-6,TNF-α和NF-κB)产生、分泌和活性。
- 雌激素与破骨细胞(骨髓中的巨噬细胞)的活性有关,绝经后的低雌激素水平降低了破骨细胞活性,使得骨骼分解大于重建速度,结果导致骨质疏松症。
- 睾丸激素水平降低与炎性标志物(IL-6和IL-6受体)增加相关。
- 保持性激素水平与降低多种炎症性疾病的风险有关,包括动脉粥样硬化、冠心病、哮喘和类风湿关节炎等。
- 吸烟和二手烟:香烟烟雾含有几种炎症诱导剂,特别是活性氧。长期吸烟增加几种促炎细胞因子(TNF-α,IL-1β,IL-6,IL-8)的产生,同时减少产生抗炎分子等。吸烟增加牙周病的风险,牙周病是增加全身炎症的独立危险因素。
- 睡眠障碍:炎症细胞因子(TNF-α和IL-1β)的产生似乎遵循昼夜节律并且可能参与动物和人类睡眠调节。睡眠失调可导致促炎分子的日间升高。包括白天嗜睡、睡眠呼吸暂停等患者的TNF-α和/或IL-6的血浆水平升高。
- 其他诱因:
- 牙周疾病:可产生全身性炎症反应,影响心脏、肾脏系统。因此,牙周病是心血管病的危险因素。
- 压力:可导致炎性细胞因子IL-6释放。压力与睡眠紊乱、体重增加有关,二者都是诱发炎症的独立因素。
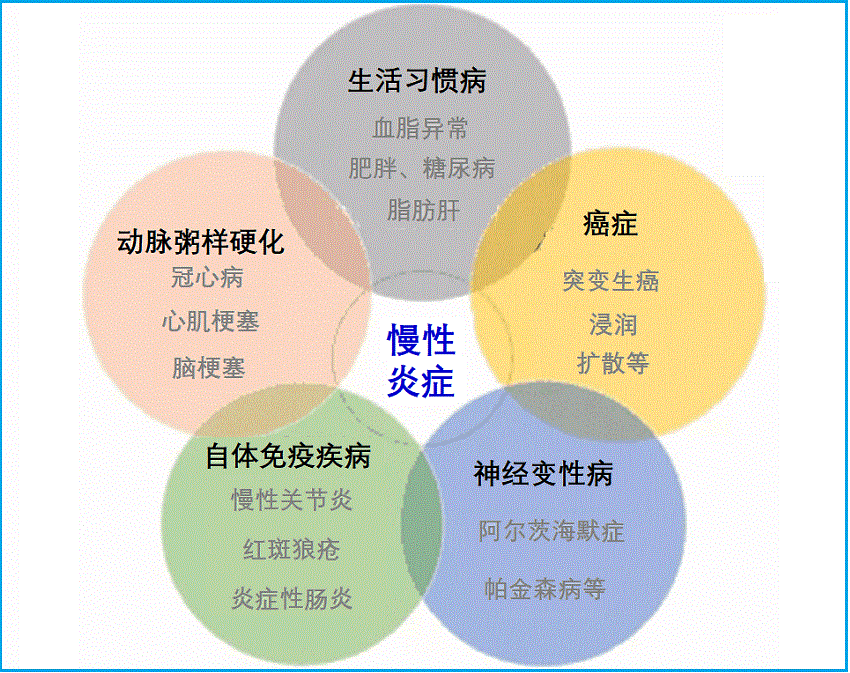
与慢性炎症相关的疾病
慢性低度炎症与许多慢性病密切相关。- 心血管疾病:炎症促成动脉粥样硬化,循环炎性因子可预测外周动脉疾病、心力衰竭、房颤、中风和冠心病等。
- 癌症:慢性低度炎症与许多类型的癌症如淋巴瘤、前列腺癌、卵巢癌、胰腺癌、结肠直肠癌和肺癌等有直接联系。有些炎症机制可能引起癌变,包括基因表达改变、DNA突变、表观遗传改变、肿瘤血管形成的促进等。
- 糖尿病:促炎细胞因子明显降低了胰岛素敏感性。II型糖尿病患者中巨噬细胞向脂肪组织的浸润及其后促炎细胞因子的释放发生率更高。
- 黄斑变性:研究表明,黄斑病变患者具有显著的高血清CRP水平等。
- 慢性肾脏病:慢性低度炎症可导致血液中几种促炎因素(包括细胞因子、AGEs和高血同)滞留、排泄减少,并可加速身体其他的慢性炎症疾病,如心脑血管病等。
- 骨质疏松症:炎性细胞因子(TNF-α,IL-1β,IL-6)参与正常骨代谢,但升高推动骨吸收,并可引起牙周病、炎性肠病和类风湿关节炎等患者骨丢失。更年期炎性因子水平增加刺激骨丢失、导致骨松症。
- 抑郁症:与升高的IL-6和CRP显著相关联,这在许多人群研究中已被观察到。
- 认知功能下降:老年人的慢性低水平炎症(如IL-6和CRP)与认知衰退和痴呆相关,包括血管性痴呆和阿尔茨海默病等。
- 自体免疫疾病:循环炎症细胞因子持续存在和升高、对抗自身组织的免疫反应即为自身免疫性疾病,如类风湿性关节炎、红斑狼疮、胰腺炎和炎症性肠炎等。
对慢性炎症的医学干预
值得注意的是,主流医学通常不会治疗慢性炎症或低度炎症,除非在炎症与某种健康状况(例如关节炎)相关的情况下进行常规医学干预。当前,常规的预防炎症的医学方法仅限于使用CRP预测高危人群的心血管疾病,以及预防性使用阿司匹林之类的药物来抑制与血栓形成相关的炎症级联反应(失控的凝血)。确实,低度炎症的潜在无症状性质使得促炎细胞因子的升高可能在一段时间内未被发现,只有在它们造成足够的细胞损伤以产生疾病症状后才被发现和治疗。- 己酮可可碱(Pentoxifylline):一种用于治疗涉及大脑、肢体和小血管等供血不足疾病的药物。己酮可可碱的强效抗炎特性是次要发现,但机制仍未完全了解。不过,研究显示己酮可可碱可调节TNF-α信号传导。在一项试验中,每天两次服用400mg可显着抑制超敏C反应蛋白(hs-CRP),慢性肾脏病患者的纤维蛋白原和TNF-α水平。
- 二甲双胍(Metformin):能量代谢和炎症的调节密切相关。代谢紊乱(肥胖症、糖尿病等)和低度炎症的共同发生可证明这一点。二甲双胍可通过增加IL-1β受体拮抗剂的产生来降低炎性细胞因子的活性,IL-1β受体拮抗剂是一种干扰IL-1β的促炎信号传导。它也可以促进良好的CRP水平。高血压和血脂异常患者服用1,700 mg/day二甲双胍12周的随机对照试验与基线水平相比,IL-6降低了26.7%,TNF-α降低了8.3%,降低程度与强效他汀类药物瑞舒伐他汀(Crestor)相似。二甲双胍的消炎作用似乎很快,在一项对128例II型糖尿病血脂异常患者进行的大型研究中,仅30天后就观察到循环TNF-α,IL-1β,CRP和纤维蛋白原的减少。
- 阿司匹林(Aspirin):在炎症分子机理发现之前就已经被用作抗炎治疗药物。它是有效的环氧合酶(COX)抑制剂。
- 阿司匹林对COX分子的修饰对心血管健康具有重要意义。血小板利用COX产生血栓素A2 - 在凝血过程的初始阶段是重要信号分子。
- 阿司匹林对血小板中COX酶的抑制作用可以部分解释其对多种疾病,包括高血压、心脏病发作和卒中等并发症的保护作用。
- 阿司匹林对COX2的抑制作用还有助于解释其对降低癌症风险的潜在作用。一些研究认为,COX-2在增加突变细胞的增殖、肿瘤形成、肿瘤侵袭和转移以及耐药性等起着一定作用。
- 他汀类药物(Low-dose Statin Drugs):研究认为,他汀类药物通过一种不同于其对胆固醇代谢的机制来减轻炎症。它们干扰白细胞表面的细胞因子受体的功能。因此,血液中的促炎信号无法刺激白细胞的反应。
关注饮食和生活方式
- 饮食直接影响慢性炎症形成和发展:
- 高血糖指数(GI)食物升高hs-CRP,饱和脂肪饮食也增加炎症的发生。这些均与冠心病、2型糖尿病和中风有关。
- 健康饮食,如丰富的蔬果、坚果和适量的鱼肉包括深海鱼类,较少量的红肉,以及泉谷类食物等,可以减少促炎因子产生,防止慢性炎症。膳食纤维(每天超过24g)摄入可以显著降低CRP,以及IL-6和TNF-α较低。
- 热量平衡:由于脂肪组织(尤其是腹部脂肪)表达炎性细胞因子,因此肥胖可能是导致轻度全身性炎症的主要原因。因此,总热量摄入应与能量消耗平衡,以免超重或肥胖。
- 运动锻炼:研究发现,运动可以降低多种促炎分子和全身性炎症分子。
- 不抽烟,或戒烟。
- 不饮酒,或适量饮酒。
- 学会压力管理,包括放松身心、良好睡眠等
慢性炎症干预路径
1.抑制炎症的基础营养:1). 维生素D:
维生素D似乎通过抑制促炎性前列腺素和抑制炎症介质NF-κB发挥抗炎活性1。尽管缺乏对其在人类中抗炎活性的干预研究,但几项观察性研究表明,维生素D缺乏可能会促进炎症。维生素D缺乏症在炎症性疾病(包括类风湿性关节炎、炎症性肠病、系统性红斑狼疮和糖尿病)患者中比在健康人中更常见2。在易发生低水平炎症的人群中也更常见3,手术后维生素D水平可能下降(一种与急性炎症相关的情况),伴随着CRP的升高4。
在一项对548名心力衰竭患者的研究中,维生素D水平低与CRP升高有关5,在一组46名患有内皮功能障碍的中年男性中,与IL-6和NF-κB的升高有关6。
2). 维生素E:
维生素E在体内起抗氧化剂的作用。具体来说,维生素E被掺入低密度脂蛋白(LDL)颗粒中,并保护它们免受氧化损伤;它似乎也通过其他机制预防动脉粥样硬化7。维生素E的γ-生育酚形式似乎补充了α-生育酚的抗炎作用。γ-生育酚已被证明可抑制COX-2并减弱IL-1β信号传导8,9。
在一项针对代谢综合征受试者的小型临床试验中,与安慰剂相比,γ-生育醇和α-生育酚的组合有效抑制了C反应蛋白和TNF-α水平10。在本研究中,两种生育酚的组合比单独使用任何一种都要好,这促使研究人员指出,“在氧化应激和炎症的生物标志物方面,α-生育酚和γ-生育酚的联合补充似乎优于单独补充,需要在前瞻性临床试验中进行测试……”。
3). 镁:
在两项大型观察研究(妇女健康倡议和哈佛护士研究)中,镁摄入量越高,hs-CRP、IL-6和TNF-α受体(衡量TNF-α活性的指标)越低11,12。来自动脉粥样硬化多民族研究的数据未能发现镁摄入量最高和最低的个体之间IL-6或CRP水平的显著差异,但确实发现饮食中较高的镁与较低水平的炎症相关蛋白同型半胱氨酸和纤维蛋白原之间存在显著关联13。
在饮食炎症指数中,镁被评为最抗炎的饮食因子,该指数根据人类和动物的实验和观察数据,对42种常见的饮食成分进行了评级,以降低CRP水平14。
4). 锌和硒:
含锌和硒的抗氧化蛋白(如超氧化物歧化酶和谷胱甘肽过氧化物酶)可减少活性氧(自由基),从而间接抑制NF-κB活性,并阻止几种炎症酶和细胞因子的产生。锌还能以更直接的方式抑制NF-κB15,16。在儿童和老年人等容易缺锌的人群中,补充锌与炎症的减少有关17,18。在几项研究中,适量补充锌可以降低老年人的低水平炎症和循环促炎因子(CRP、TNF-α、IL-6和IL-8)得到了降低19-21。与锌一样,硒缺乏在与疾病相关的慢性炎症状态(如败血症)中很常见22,144补充硒与减少炎症和改善患者预后有关16。
5).鱼油:
鱼油是ω-3脂肪酸二十碳五烯酸(EPA)和二十二碳六烯酸(DHA)的最佳来源,这些脂肪酸只能在人类中有限程度地合成。Omega-3脂肪酸在预防数万名患者的心血管疾病和死亡率方面已经得到了很好的研究;ω-3的抗炎作用有助于这种活性23。在许多其他炎症性疾病的研究中,特别是哮喘、炎症性肠病和类风湿性关节炎,ω-3也被证明能成功地改善患者的预后24,25。
几项大型观察性试验的数据证实了鱼油/ω-3摄入量增加与全身炎症减少之间的联系。在来自卫生专业人员随访研究的855名健康参与者中,ω-3脂肪酸的摄入与血浆TNF-α活性标志物水平较低有关;有趣的是,高摄入ω-3和ω-6脂肪酸(通常被认为是促炎性的)与炎症水平最低有关26。由727名女性组成的护士健康研究I队列显示,在摄入ω-3最多的20%人群中,炎症标志物(包括CRP和IL-6)的浓度较低27。在ATTICA对3000多名没有任何心血管疾病证据的希腊男性和女性进行的研究中,每周食用300g以上鱼类的参与者平均CRP降低33%,IL-6降低33%,并且TNF-α比不吃鱼的参与者低21%28。在来自动脉粥样硬化多民族研究(MESA)队列的5677名没有心血管疾病的男性和女性样本中,长链ω-3的摄入(来自鱼类或补充剂)与多种炎症标志物(包括CRP、IL-6和TNF-α受体,一种衡量TNF-α活性的指标)的血浆浓度降低有关29。
6). 特异性促分解介质(SPMs):
特异性促分解介质(SPMs)是来源于多不饱和脂肪酸代谢的细胞信号分子。它们在解决炎症反应中起着重要作用30,31。SPMs包括脂氧素(lipoxins)、消炎素(Resolvins)、保护素(Protectins)和马雷素(Maresins)及其前体物。它们不仅抑制炎症,而且积极解决炎症过程,帮助组织恢复正常状态31。SPMs招募巨噬细胞清除细胞碎片,从而恢复健康细胞32;使炎症损伤的组织恢复活力33;并恢复抗炎细胞因子和促炎细胞因子之间的平衡34,35。
慢性炎症会导致衰老和变性,SPMs可能有助于降低与炎症和衰老相关的某些疾病的风险31,36-39。对肥胖小鼠的研究表明,低度炎症会导致代谢紊乱,SPMs前体可以减轻肥胖并发症38。另一项研究表明,在喂食高胆固醇饮食九周的小鼠中,每天1-5mg/kg的SPMs Resolvin E1可以减少动脉粥样硬化病变37。
源自深海鱼油的SPMs有助于支持身体有效解决炎症反应。身体从ω-3脂肪酸中产生SPMs,包括EPA和DHA40;然而,它们并没有完全转化为SPMs。Omega-3和SPMs在体内也有不同的作用。
为了更好地了解补充鱼油(EPA和DHA)对SPMs血浆脂质代谢物浓度的影响,进行了一项新的临床试验。共有21名患有慢性炎症的患者被随机分组,依次接受3g/天的EPA或DHA,为期10周;其间有10周的洗脱期。这项研究还包括一项离体分析,评估了EPA和DHA衍生的SPMs对炎症细胞(单核细胞)的影响有何不同41。
结果表明,EPA和DHA都以各自的方式提高了血浆中SPMs和SPM前体的水平。在研究的离体部分,从参与者的血液样本中提取的单核细胞(巨噬细胞前体细胞)在体外用毒素(LPS)刺激以诱导炎症反应。EPA和DHA在体外均能降低促炎因子TNF-α和单核细胞趋化蛋白-1(MCP-1)的基因表达。EPA对促炎IL-6的表达没有影响;然而,DHA降低了它。DHA降低了抗炎IL-10的表达;然而,EPA却增加了它。有趣的是,DHA比EPA更有效地抑制单个的促炎细胞因子,但EPA更有效地平衡炎症细胞因子与抗炎细胞因子IL-10的比例。
这些结果为EPA和DHA的独立作用提供了新的见解,并表明这两种ω-3脂肪酸在炎症缓解中发挥着重要作用。
2. 抑制炎症的功能性营养:
1).脱氢表雄酮(DHEA):
性激素水平低与炎症标记物的系统性增加有关42。作为一种肾上腺类固醇激素,DHEA是性类固醇睾酮和雌激素的前体。DHEA在年轻人中含量丰富,但随着年龄的增长而稳步下降,可能是与年龄相关的性类固醇减少的部分原因43。在细胞培养和动物模型中,DHEA可以抑制炎症细胞因子活性,在某些情况下比睾酮或雌激素更有效44。慢性炎症本身可能会降低DHEA水平45。在一项小型研究中,老年志愿者补充DHEA(50mg/天,持续两年)可显著降低TNF-α和IL-6水平,降低内脏脂肪量,改善糖耐量(二者均与炎症有关)46。
2).姜黄素:
广泛的体外和动物研究已经检验了姜黄素对实验诱导的炎症性疾病(动脉粥样硬化、关节炎、糖尿病、肝病、胃肠道疾病和癌症)和疾病标志物(脂氧合酶、环氧合酶、TNF-α、IL-1β、NF-κB等)的影响47,48。研究姜黄素对炎症性疾病患者预后影响的人类研究较少,但大多数姜黄素的小型随机对照试验始终显示,患者在几种炎症性疾病方面有所改善,包括银屑病、肠易激综合征(IBS)、类风湿性关节炎和炎症性眼病49,50。
3).白藜芦醇和紫檀芪:
白藜芦醇发挥抗炎活性的确切机制尚未确定,尽管它在动物模型和人类细胞培养中抑制多种促炎化合物(环加氧酶、TNF-α、IL-1β、IL-6、NF-κB)51,52。其相关化合物紫檀芪在细胞培养中显示出类似的炎症标志物抑制作用53。
炎症免疫反应的调节可能有助于白藜芦醇在心脏病、癌症、急性胰腺炎和炎症性肠病动物模型中的保护作用54。白藜芦醇也可以预防一般的、低水平的副炎症:在对10名健康志愿者进行的一项小型交叉研究中,当与一顿高脂肪、高碳水化合物(930千卡)一起服用时,白藜芦醇(100mg)可以防止餐后氧化和炎症标志物的急剧增加。例如,在试验餐后5小时内,IL-1β的合成增加了91%;使用白藜芦醇时,这种增加明显较少(29%)55。
4).茶多酚:
数十项体外和动物研究证实了绿茶和红茶多酚的抗炎作用56。绿茶多酚EGCG和茶黄素通过抑制NF-κB信号通路发挥抗炎作用,它降低了细胞培养实验中几种炎症蛋白(脂氧合酶、环氧合酶、TNF-α、IL-1β、IL-6和IL-8)的表达57。EGCG还抑制组胺的产生和释放,组胺是过敏和炎症反应的关键介质58。
在喝茶的观察性研究中,与不喝茶的人相比,每天喝2杯茶(红茶或绿茶)可使CRP降低近20%,并显著降低其他两种炎症标志物(血清淀粉样蛋白a和半抗原,在冠心病中升高)的水平59。在临床干预中,红茶在降低炎症标志物方面似乎比绿茶更成功11。117在一项小型试验中,健康的非吸烟者服用红茶提取物(相当于每天四杯茶)六周,CRP也降低了25%60。在一项大型研究中,对患有冠心病的高风险健康人也观察到了类似的平均降低,但在CRP起始值最高(>3 mg/L)的个体中,CRP降低了40-50%61。
5). 乳香:
乳香是阿育吠陀医学中一种传统的抗关节炎药物;其抗炎特性被归因于对5-LOX的特异性抑制和乳香酸(乳香主要活性成分)减少促炎白三烯的产生62。在细胞培养中,粗提物和高度纯化的乳香提取物都抑制了促炎TNF-α和IL-1β的产生63。其中一种乳香酸乙酰-11-酮- β -乳香酸(AKBA)在小鼠中是NF-κB活性的抑制剂64;而在啮齿动物炎症模型中,四种最丰富的乳香酸的局部混合物可以减轻炎症65。
最近对乳香治疗炎症条件的人体试验的系统综述表明,对该提取物的少量随机对照试验为其用于哮喘和骨关节炎产生了令人鼓舞的结果66。因此需要进行更大规模的研究来证实提取物是一种有效的治疗方法。标准化乳香提取物(30%AKBA)在减轻骨关节炎患者的疼痛方面是有效的67。当与非挥发性乳香精油结合使用时,标准化提取物(AprèsFlex或Aflapin)在较低浓度下表现出更高的活性68。尽管结果不一,但乳香提取物用于炎症性肠病的用途已在多项临床试验中进行了研究69-71。
6). 类胡萝卜素:
在妇女健康和衰老研究中,血液中α-胡萝卜素和总类胡萝卜素水平最高的参与者在研究开始时比类胡萝卜素水平较低的参与者有更低的IL-6水平72。血液中α-和β-胡萝卜素、叶黄素/玉米黄质或总类胡萝卜素水平最低的参与者在2年的时间里更有可能经历IL-6的增加。
7). N-乙酰半胱氨酸(NAC):
NF-κB通路的激活在炎症细胞因子基因的激活中起着核心作用;NAC在细胞培养中抑制NF-κB,降低IL-6和IL-8等细胞因子的表达73,74。确定NAC在降低人类慢性炎症方面的作用的数据有限,但显示出前景。在慢性肾脏病患者中,补充NAC八周后,循环IL-6水平略有下降,但具有统计学意义75。研究开始时,对有严重炎症的人的影响更为明显(通过hs-CRP测量)。在一项针对烧伤患者的小型研究中,NAC还降低了全身炎症标志物76。
8). 菠萝蛋白酶:
菠萝蛋白酶是来源于菠萝茎的包括蛋白水解酶的混合物。在临床前研究中,菠萝蛋白酶一直表现出多种抗炎特性77,78。这些抗炎作用归因于COX-2活性降低和前列腺素合成减少。菠萝蛋白酶还通过对纤维蛋白和纤维蛋白原的作用调节凝血78。
菠萝蛋白酶治疗炎症条件的人体试验也产生了有希望的结果79。一项对四项研究的荟萃分析发现,菠萝蛋白酶在牙科手术后可以减轻肿胀,并具有镇痛和抗炎作用80。
一项为期一个月的开放标签研究评估了不同剂量菠萝蛋白酶对轻度急性膝关节疼痛的影响。77名膝盖疼痛不到三个月但身体健康的参与者每天服用200或400mg菠萝蛋白酶。参与者在基线和一个月后通过标准化问卷评估自己的疼痛和功能。与基线相比,200mg组的症状评分改善了41%,400mg组改善了59%。两组患者的心理健康状况也显著改善81。
一项随机对照非劣效性试验在103名膝骨关节炎患者中比较了含有菠萝蛋白酶(90mg,每日三次)加胰蛋白酶和芸香苷的标准化酶制剂与非甾体抗炎药双氯芬酸(Voltaren等)(50mg,每日二次)。研究人员发现,菠萝蛋白酶在患者报告的疼痛和功能评估中的作用至少与双氯芬酸一样好80。
另一项随机试验招募了30名有症状的颞下颌关节炎患者,比较了三种治疗方案:
- 单独使用双氯芬酸(第1组)。
- 双氯芬酸加上由菠萝蛋白酶(90mg)、胰蛋白酶和芸香糖苷组成的酶制剂(第2组)。
- 单独的酶制剂(第3组)。
第1组和第3组的反应被认为是相同的,再次证明含有菠萝蛋白酶的组合与双氯芬酸一样有效。然而,与单独使用双氯芬酸或口服酶治疗的参与者相比,第2组的参与者在疼痛和炎症方面有显著改善82。这些发现表明菠萝蛋白酶和这些其他成分可以补充非甾体抗炎药在炎症条件的管理。
一项针对49名骨关节炎患者的试点研究发现,食用含有乳香和菠萝蛋白酶混合物的补充剂长达六个月,可以显著提高自我报告的生活质量分数。据报道,基线时受影响最严重的关节症状改善最为明显83。
在另一项随机对照非劣效性试验中,研究人员将90名患有疼痛性髋关节骨关节炎的患者分为两组:一半接受含有菠萝蛋白酶、胰蛋白酶和芸香苷的口服酶制剂治疗六周,另一半接受抗炎药双氯芬酸治疗。他们发现,菠萝蛋白酶制剂在疼痛、僵硬和身体功能的标准化评分中与双氯芬酸一样有效,而且往往比药物对照品耐受性更好。此外,研究人员报告称,71%服用含菠萝蛋白酶制剂的受试者和61%服用双氯芬酸的受试人员的疗效为“良好”或“非常好”77,78。
菠萝蛋白酶与歌手等声带炎症:
菠萝蛋白酶等纯天然的抗炎药可能对歌手经常出现的声带炎症有用。
歌手、教师和其他经常使用或过度使用声音的人(有时被称为误用甚至滥用)有很高的声带紧张风险,这可能会导致炎症和肿胀84-86。这种慢性刺激会导致息肉和结节的发展87,88。这些问题通常可以通过休息声音(如果可能的话)、正确使用声音、口腔补水和使用室内加湿器来解决;在某些情况下,可能需要口服或注射皮质类固醇,甚至手术切除84-86,88,89。当导致病变发展的活动(如唱歌或大喊大叫)持续时,这些病变复发并不罕见。因此,经常寻求非侵入性的自然支持。
根据支持菠萝蛋白酶减轻炎症能力的证据,菠萝蛋白酶作为治疗声带肿胀和炎症的药物在歌手和声乐家中广受欢迎90。考虑到非甾体抗炎药是导致声带问题常见原因的风险因素,这一点尤为重要和有价值88,91。
一项临床研究对56名因良性声带疾病接受手术的患者进行了检查,发现术后使用菠萝蛋白酶(80mg)和乳香(750mg)联合治疗,每天两次,持续一周,可有效促进组织修复和嗓音恢复。与吸入类固醇治疗相比,这种治疗具有良好的耐受性,产生的副作用更少87。
9). 芝麻木脂素:
观察到芝麻油可以减少真菌和大鼠肝细胞中花生四烯酸的产生,从而鉴定出芝麻木脂素(芝麻脂素/Sesamin、芝麻林素/Sesamolin、芝麻明酚/Sesaminol)是合成花生四烯酸所使用的酶之一Δ5去饱和酶(δ -5去饱和酶)的特异性抑制剂92。通过抑制Δ5去饱和酶,芝麻木脂素可以减少促炎前列腺素、白三烯和凝血素的合成,这些都需要花生四烯酸作为起始物质93。
在动物模型中,高芝麻油的饮食减少了促炎前列腺素PGE-1和-2以及血栓素B2的产生94。在人类中,五周的芝麻素补充(39mg/天)减少了促炎血管收缩剂20-羟基二十碳四烯酸(20-HETE,酶5-LOX的产物)减少30%95。芝麻木脂素的这种潜在抗炎特性可能部分解释了其观察到的降压活性96。
10).黑籽油:
黑籽油(或称黑孜然籽油)是一种抗氧化剂和抗炎剂,传统上用于促进消化、皮肤和肝脏健康。261研究表明,黑籽油平衡免疫和炎症反应,支持健康的免疫系统功能97,98。随着年龄的增长,破坏性炎症途径增加、免疫反应受损,这与关节炎、过敏和心血管疾病等慢性疾病有关99。
黑籽油及其活性成分百里香醌(Thymoquinone)被认为可以抑制有害炎症,同时支持抵抗感染和癌症所需的正常免疫反应98,100。黑籽油可刺激巨噬细胞和辅助T细胞的活性101。
在一项安慰剂对照试验中,40名患有类风湿性关节炎的女性每天服用安慰剂一个月,然后每天两次服用500mg黑籽油、持续一个月。据报道,关节炎症状,包括关节肿胀和晨僵,治疗后疾病严重程度有所下降102。在一项前瞻性双盲研究中,66名过敏性鼻炎患者暴露于黑孜然籽油中30天。结果鼻炎的症状,包括充血、鼻子发痒和打喷嚏,在治疗后有所减轻103。动物研究表明,补充黑籽油具有心脏保护作用,因为它可以减少脂质氧化,促进健康的胆固醇水平104。
3. 支持线粒体:
线粒体呼吸过程中产生的活性氧会导致炎症,如上述有关内容所述。衰老个体特别容易受到线粒体相关氧化应激的影响,因为线粒体随着年龄的增长而变得越来越功能障碍。采取措施支持线粒体的完整性和效率可以帮助减轻线粒体功能不良引起的一些系统氧化和炎症负担。辅酶Q10和吡咯喹啉醌(PQQ)这两种营养素是强大的线粒体保护剂,研究支持这些化合物的抗炎作用105,106。
1).吡咯喹啉醌(PQQ):
PQQ是对细胞能量稳态和氧化还原平衡至关重要的酶的辅因子107。多项研究表明,PQQ在线粒体环境应激和氧化负荷增加期间发挥保护作用106,108。在一项研究中,给予补充PQQ的饮食的大鼠表现出更大的能量消耗,肝组织线粒体密度增加。补充PQQ的大鼠甘油三酯也较低,与未服用PQQ的鼠相比,它们的心脏更能防止缺氧109。在心脏组织氧气供应有限的时期,氧化应激和随后的炎症损伤细胞急剧上升;该动物模型的研究表明,PQQ可以通过在不利条件下保持线粒体效率来避免这种炎症细胞破坏。
2). 辅酶Q10:
CoQ10是线粒体ATP产生过程中不可或缺的中间产物。研究表明,在炎症条件下,辅酶Q10水平较低。在一项研究中,感染性休克患者的辅酶Q10水平明显低于健康人,并且在患者中,较低的CoQ10水平与较高水平的一种炎症介质VCAM相关110。
在动物模型中,给大鼠饮用添加果糖的饮用水,该实验会导致肥胖、糖尿病和其他炎症并发症。补充辅酶Q10可通过降低肝脏中CRP和其他炎症介质的表达来减轻炎症反应111。实验室研究表明,辅酶Q10调节数百个基因的表达,其中许多与炎症信号有关112。特别重要的是,一项实验表明,在生理相关浓度下,CoQ10能够通过调节NF-κB信号通路,将诱导的TNF-α减弱25%以上112。
4. 预防炎症性糖化反应:
已知升高的血糖和糖基化终产物(AGEs)在引发炎症风暴中的作用。幸运的是,除了减少热量摄入以抑制空腹和餐后葡萄糖浓度外,一些天然化合物还能改善糖基化过程,并可能有助于控制糖诱导的炎症级联反应。在这些抗糖基化营养素中,最主要的是B族维生素家族成员苯磷硫胺和一种氨基酸肌肽。
1). 苯磷硫胺:
自20世纪90年代中期以来,苯磷硫胺一直被用于治疗糖尿病并发症113。最近的证据继续支持它作为一种强大的保护剂来对抗血糖诱导的组织损伤。在一项临床试验中,165名糖尿病受试者被随机分配接受每天300或600mg苯磷硫胺或安慰剂治疗六周。干预期结束后,服用苯磷硫胺的患者表现出剂量依赖性的神经性疼痛改善114。一个动物模型发现,苯磷硫胺通过有效抑制炎症来缓解神经性疼痛115。此外,实验室研究表明,除了阻断糖基化反应外,苯磷硫胺可能通过调节COX和LOX酶活性更直接地调节炎症116。
2). 肌肽:
肌肽在体内发挥一系列有利的生物化学作用。它能有力地减缓糖基化反应并缓解氧化应激117。此外,几项实验表明,肌肽具有显著的抑制各种细胞类型炎症的能力118-120。不幸的是,肌肽水平在10岁至70岁之间下降了63%121。此外,在2型糖尿病患者中,骨骼肌肌肽含量明显低于健康对照受试者122。当肌肽作为化学诱导糖尿病动物的补充剂时,它能够保护脆弱的视网膜细胞免受与高血糖相关的炎症并发症的影响123。
3). 绞股蓝:
绞股蓝在亚洲医学中用于治疗多种健康状况,包括血脂异常、2型糖尿病和炎症124。它的作用至少部分是由于它能够激活一种单磷酸腺苷活化蛋白激酶(AMPK)的关键酶。这种影响葡萄糖代谢和脂肪储存的酶被称为“代谢主开关”,因为它控制着从食物中提取能量并在全身储存和分配能量的许多途径125。
超重对AMPK激活和慢性炎症有显著影响;它抑制AMPK的激活,导致腹部脂肪沉积,进而激活全身炎症。同时,炎症本身抑制AMPK的激活,产生粘性循环126。
然而,更多的AMPK激活有助于减肥并可以抑制炎症127-129。此外,AMPK激活增加与肝脏脂肪积聚减少有关,这是炎症化学物质的另一个来源130。
有证据表明绞股蓝具有抗炎作用。在一项实验室研究中,研究人员发现该草药提取物能显著抑制几种炎症化学物质,包括TNF-α、白细胞介素-6和COX-2信使核糖核酸(mRNA)131。另一项针对24名2型糖尿病患者的研究发现,饮用该草药泡茶12周可显著降低胰岛素抵抗,而胰岛素抵抗是导致全身炎症的关键因素132,133。
4).橙皮苷:
橙皮苷和相关黄酮类化合物存在于多种植物中,尤其是柑橘类水果的果皮中134,135。橙皮苷的消化会产生一种称为陈皮素(Hesperetin)的化合物以及其他代谢产物。这些化合物是强大的自由基清除剂,具有抗炎、胰岛素增敏和降脂活性136,137。动物和体外研究结果表明,橙皮苷对血糖和脂质水平的积极作用可能部分与AMPK途径的激活有关138-140。越来越多的证据表明,橙皮苷可能有助于预防和治疗与衰老相关的许多慢性疾病136。
橙皮苷可以部分通过激活AMPK信号通路来预防糖尿病及其并发症。巧合的是,二甲双胍,一种领先的糖尿病药物,也激活AMPK通路。在一项针对24名糖尿病参与者的为期六周的随机对照试验中,每天补充500mg橙皮苷可以改善血糖控制,提高总抗氧化能力,减少氧化应激和DNA损伤141。另一组研究人员使用尿陈皮素作为膳食橙皮苷的标志物,发现摄入橙皮苷水平最高的人在4.6年内患糖尿病的风险比摄入橙皮苷水平最低的人低32%142。
在一项随机对照试验中,24名患有代谢综合征的成年人每天服用500mg橙皮苷或安慰剂治疗三周。洗脱期后,用橙皮苷重复试验,安慰剂分配颠倒。橙皮苷治疗改善了内皮功能,这可能是其有益于心血管系统的重要机制之一。相对于安慰剂,补充橙皮苷还导致炎症标志物hs-CRP的中位水平降低33%,总胆固醇、载脂蛋白B(apoB)和血管炎症标志物的水平显著降低139。在另一项针对有证据表明存在血管功能障碍的超重成年人的随机对照试验中,连续六周每天补充450mg橙皮苷可降低血压,降低血管炎症标志物143。另一项对照临床试验包括75名心脏病发作患者,他们被随机分配接受每天600mg橙皮苷或安慰剂治疗四周。服用橙皮苷的患者的高密度脂蛋白胆固醇、血管炎症标志物、脂肪酸和葡萄糖代谢水平显著改善144。
综合干预方案
据于上述慢性炎症干预路径及相关营养素或功能营养,其综合干预方案如下:- 慢性炎症管理要略:
- 慢性炎症管理:
以及参阅本网站如下专文了解更多内容:
参考文献:
1. Krishnan AV et al. The role of vitamin D in cancer prevention and treatment. Endocrinol Metab Clin North Am. 2010;39(2):401–18
2. Guillot X et al. Vitamin D and inflammation. Joint Bone Spine. 2010;77(6):552–557
3. Awad AB et al. Vitamin D and metabolic syndrome risk factors: evidence and mechanisms. Crit Rev Food Sci Nutr. 2012;52(2):103–112
4. Reid D et al. The relation between acute changes in the systemic inflammatory response and plasma 25-hydroxyvitamin D concentrations after elective knee arthroplasty. American Journal of Clinical Nutrition. 2011;93(5):1006–1011
5. Liu LCY et al. Vitamin D status and outcomes in heart failure patients. Eur. J. Heart Fail. 2011;13(6):619–625
6. Jablonski KL et al. 25-Hydroxyvitamin D deficiency is associated with inflammation-linked vascular endothelial dysfunction in middle-aged and older adults. Hypertension. 2011;57(1):63–69
7. Meydani M. Vitamin E and atherosclerosis: beyond prevention of LDL oxidation. J Nutr. 2001Feb;131(2):366S-8S.
8. Jiang Q et al. gamma-tocopherol and its major metabolite, in contrast to alpha-tocopherol, inhibit cyclooxygenase activity in macrophages and epithelial cells. Proc Natl Acad Sci USA. 2000 Oct 10;97(21):11494-9.
9. Sjoholm A et al. gamma-tocopherol partially protects insulin-secreting cells against functional inhibition by nitric oxide. Biochem Biophys Res Commun. 2000 Oct 22;277(2):334-40.
10. Devaraj S et al. Gamma-tocopherol supplementation alone and in combination with alpha-tocopherol alters biomarkers of oxidative stress and inflammation in subjects with metabolic syndrome. Free Radic Biol Med. 2008;44(6):1203-8.
11. Galland L. Diet and inflammation. Nutr Clin Pract. 2010;25(6):634–640
12. Chacko S et al. Relations of dietary magnesium intake to biomarkers of inflammation and endothelial dysfunction in an ethnically diverse cohort of postmenopausal women. Diabetes. 2010;
13. de Oliveira Otto et al. Dietary micronutrient intakes are associated with markers of inflammation but not with markers of subclinical atherosclerosis. Journal of Nutrition. 2011;141(8):1508–1515
14. Cavicchia PP et al. A new dietary inflammatory index predicts interval changes in serum high-sensitivity C-reactive protein. Journal of Nutrition. 2009;139(12):2365–2372
15. Prasad AS. Zinc: role in immunity, oxidative stress and chronic inflammation. Current Opinion in Clinical Nutrition and Metabolic Care. 2009;12(6):646–652
16. Duntas LH. Selenium and inflammation: underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009;41(6):443–447
17. Kelishadi R et al. Effect of zinc supplementation on markers of insulin resistance, oxidative stress, and inflammation among prepubescent children with metabolic syndrome. Metab Syndr Relat Disord. 2010;8(6):505–510
18. Wong CP et al. Zinc and its role in age-related inflammation and immune dysfunction. Mol. Nutr. Food Res. 2011;
19. Bao B et al. Zinc decreases C-reactive protein, lipid peroxidation, and inflammatory cytokines in elderly subjects: a potential implication of zinc as an atheroprotective agent. American Journal of Clinical Nutrition. 2010;91(6):1634–1641
20. Kahmann L et al. Zinc supplementation in the elderly reduces spontaneous inflammatory cytokine release and restores T cell functions. Rejuvenation Res. 2008;11(1):227–237
21. Mariani E et al. Simultaneous evaluation of circulating chemokine and cytokine profiles in elderly subjects by multiplex technology: relationship with zinc status. Biogerontology. 2006;7(5-6):449–459
22. Maehira F et al. Alterations of serum selenium concentrations in the acutephase of pathological conditions. Clin. Chim. Acta. 2002;316(1-2):137–146
23. Marik PE et al. Omega-3 dietary supplements and the risk of cardiovascular events: a systematic review. Clin Cardiol. 2009;32(7):365–372
24. Calder PC. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am J Clin Nutr. 2006;83(6 Suppl):1505S–1519S
25. Giugliano D et al. The Effects of Diet on Inflammation. J Am Coll Cardiol. 2006;48(4):677–685
26. Pischon T et al. Habitual dietary intake of n-3 and n-6 fatty acids in relation to inflammatory markers among US men and women. Circulation. 2003;108(2):155–160
27. Lopez-Garcia E et al. Consumption of (n-3) fatty acids is related to plasma biomarkers of inflammation and endothelial activation in women. J Nutr. 2004;134(7):1806–1811
28. Zampelas A et al. Fish consumption among healthy adults is associated with decreased levels of inflammatory markers related to cardiovascular disease: the ATTICA study. J Am Coll Cardiol. 2005;46(1):120–124.
29. He K et al. Associations of dietary long-chain n-3 polyunsaturated fatty acids and fish with biomarkers of inflammation and endothelial activation (from the Multi-Ethnic Study of Atherosclerosis [MESA]). Am J Cardiol. 2009;103(9):1238–1243
30. Serhan CN et. Novel anti-inflammatory--pro-resolving mediators and their receptors. Current topics in medicinal chemistry. 2011;11(6):629-647.
31. Basil MC et al. Specialized pro-resolving mediators: endogenous regulators of infection and inflammation. Nature Reviews Immunology. 2016;16(1):51-67.
32. Serhan CN. Treating inflammation and infection in the 21st century: new hints from decoding resolution mediators and mechanisms. Faseb j. 2017;31(4):1273-1288.
33. Hasturk H et al. Resolvin E1 regulates inflammation at the cellular and tissue level and restores tissue homeostasis in vivo. J Immunol. 2007;179(10):7021-7029.
34. Serhan CN. Pro-resolving lipid mediators are leads for resolution physiology. Nature. 2014;510(7503):92-101.
35. Serhan CN. Discovery of specialized pro-resolving mediators marks the dawn of resolution physiology and pharmacology. Mol Aspects Med. 2017;58:1-11.
36. Wang X et al. Insufficient resolution response in the hippocampus of a senescence-accelerated mouse model--SAMP8. J Mol Neurosci. 2015;55(2):396-405.
37. Salic K et al. Resolvin E1 attenuates atherosclerosis in absence of cholesterol-lowering effects and on top of atorvastatin. Atherosclerosis. 2016;250:158-165.
38. Neuhofer A et al. Impaired local production of proresolving lipid mediators in obesity and 17-HDHA as a potential treatment for obesity-associated inflammation. Diabetes. 2013;62(6):1945-1956.
39. Huang J et al. Targeting the D Series Resolvin Receptor System for the Treatment of Osteoarthritis Pain. Arthritis Rheumatol. 2017;69(5):996-1008.
40. Norris PC et al. Identification of specialized pro-resolving mediator clusters from healthy adults after intravenous low-dose endotoxin and omega-3 supplementation: a methodological validation. Scientific Reports. 2018;8(1):18050.
41. So J et al. EPA and DHA differentially modulate monocyte inflammatory response in subjects with chronic inflammation in part via plasma specialized pro-resolving lipid mediators: A randomized, double-blind, crossover study. Atherosclerosis. 2020;316:90-98.
42. Singh T et al. Inflammatory markers in population studies of aging. Ageing Res Rev. 2011;10(3):319–329
43. Heffner KL. Neuroendocrine effects of stress on immunity in the elderly: implications for inflammatory disease. Immunol Allergy Clin North Am. 2011;31(1):95–108
44. Gordon CM et al. Adrenal and gonadal steroids inhibit IL-6 secretion by human marrow cells. Cytokine. 2001;16(5):178–186
45. Ernestam et al. Increased DHEAS levels in patients with rheumatoid arthritis after treatment with tumor necrosis factor antagonists: evidence for improved adrenal function. The Journal of Rheumatology. 2007;34(7):1451–1458
46. Weiss EP et al. Dehydroepiandrosterone (DHEA) replacement decreases insulin resistance and lowers inflammatory cytokines in aging humans. Aging. 2011;3(5):533–542
47. Chainani-Wu N. Safety and anti-inflammatory activity of curcumin: a component of turmeric (Curcuma longa). J Altern Complement Med. 2003;9(1):161–168
48. Bengmark S. Curcumin, an atoxic antioxidant and natural NFkappaB, cyclooxygenase-2, lipooxygenase, and inducible nitric oxide synthase inhibitor: a shield against acute and chronic diseases. JPEN J Parenter Enteral Nutr. 2006;30(1):45–51
49. Epstein J et al. Curcumin suppresses p38 mitogen-activated protein kinase activation, reduces IL-1beta and matrix metalloproteinase-3 and enhances IL-10 in the mucosa of children and adults with inflammatory bowel disease. Br J Nutr. 2010;103(6):824–832
50. White B et al. Clinical Inquiry. Does turmeric relieve inflammatory conditions? J Fam Pract. 2011;60(3):155–156
51. Jha RK et al. Emerging role of resveratrol in the treatment of severe acute pancreatitis. Front Biosci (Schol Ed). 2010;2:168–175
52. Khanduja KL et al. Resveratrol inhibits N-nitrosodiethylamine-induced ornithine decarboxylase and cyclooxygenase in mice. J. Nutr. Sci. Vitaminol. 2004;50(1):61–65
53. Pan Z et al. Identification of molecular pathways affected by pterostilbene, a natural dimethylether analog of resveratrol. BMC Med Genomics. 2008;1:7
54. Clarke JQ et al. A review of complementary and alternative approaches to immunomodulation. Nutrition in Clinical Practice. 2008;23(1):49–62
55. Ghanim H et al. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J Clin Endocrinol Metab. 2011;96(5):1409–1414
56. Singh R et al. Green tea polyphenol epigallocatechin-3-gallate: inflammation and arthritis. [corrected]. Life Sci. 2010;86(25-26):907–918
57. de Mejia et al. Bioactive components of tea: cancer, inflammation and behavior. Brain Behav. Immun. 2009;23(6):721–731
58. Melgarejo E et al. Targeting of histamine producing cells by EGCG: a green dart against inflammation? J. Physiol. Biochem. 2010;66(3):265–270
59. De Bacquer D et al. Epidemiological evidence for an association between habitual tea consumption and markers of chronic inflammation. Atherosclerosis. 2006;189(2):428–435
60. Steptoe A et al. The effects of chronic tea intake on platelet activation and inflammation: a double-blind placebo controlled trial. Atherosclerosis. 2007;193(2):277–282
61. Bahorun T et al. Black tea reduces uric acid and C-reactive protein levels in humans susceptible to cardiovascular diseases. Toxicology. 2010;278(1):68–74
62. Boswellia serrata. Altern Med Rev. 2008;13(2):165–167
63. Gayathri B et al. Pure compound from Boswellia serrata extract exhibits anti-inflammatory property in human PBMCs and mouse macrophages through inhibition of TNFalpha, IL-1beta, NO and MAP kinases. International Immunopharmacology. 2007;7(4):473–482
64. Cuaz-Pérolin C et al. Antiinflammatory and antiatherogenic effects of the NF-kappaB inhibitor acetyl-11-keto-beta-boswellic acid in LPS-challenged ApoE-/- mice. Arteriosclerosis, Thrombosis, and Vascular Biology. 2008;28(2):272–277
65. Singh S et al. Boswellic acids: A leukotriene inhibitor also effective through topical application in inflammatory disorders. Phytomedicine. 2008;15(6-7):400–407
66. Ernst E. Frankincense: systematic review. BMJ. 2008;337:a2813
67. Sengupta K et al. A double blind, randomized, placebo controlled study of the efficacy and safety of 5-Loxin for treatment of osteoarthritis of the knee. Arthritis Res. Ther. 2008;10(4):R85
68. Sengupta K et al. Comparative efficacy and tolerability of 5-Loxin and AflapinAgainst osteoarthritis of the knee: a double blind, randomized, placebo controlled clinical study. Int J Med Sci. 2010;7(6):366–377
69. Gupta I et al. Effects of Boswellia serrata gum resin in patients with ulcerative colitis. Eur. J. Med. Res. 1997;2(1):37–43
70. Gupta I et al. Effects of gum resin of Boswellia serrata in patients with chronic colitis. Planta Med. 2001;67(5):391–395
71. Holtmeier W et al. Randomized, placebo-controlled, double-blind trial of Boswellia serrata in maintaining remission of Crohn's disease: good safety profile but lack of efficacy. Inflamm. Bowel Dis. 2011;17(2):573–582
72. Walston J et al. Serum antioxidants, inflammation, and total mortality in older women. Am. J. Epidemiol. 2006;163(1):18–26
73. Araki et al. N-acetylcysteine inhibits induction of nitric oxide synthase in 3T3-L1 adipocytes. J. UOEH. 2007;29(4):417–429
74. Radomska-Leśniewska et al. Influence of N-acetylcysteine on ICAM-1 expression and IL-8 release from endothelial and epithelial cells. J. Physiol. Pharmacol. 2006;57 Suppl 4:325–334
75. Nascimento MM et al. Effect of oral N-acetylcysteine treatment on plasma inflammatory and oxidative stress markers in peritoneal dialysis patients: a placebo-controlled study. Perit Dial Int. 2010;30(3):336–342
76. Csontos C et al. Effect of N-acetylcysteine treatment on the expression of leukocyte surface markers after burn injury. Burns. 2011;37(3):453–464
77. de Lencastre Novaes LC et al. Stability, purification, and applications of bromelain: A review. Biotechnol Prog. Jan-Feb 2016;32(1):5-13.
78. Chakraborty AJ et al. Bromelain a Potential Bioactive Compound: A Comprehensive Overview from a Pharmacological Perspective. Life (Basel). Apr 6 2021;11(4)
79. Colletti A et al. Recent Advances and Insights into Bromelain Processing, Pharmacokinetics and Therapeutic Uses. Applied Sciences. 2021;11(18):8428.
80. de Souza GM et al. Is bromelain effective in controlling the inflammatory parameters of pain, edema, and trismus after lower third molar surgery? A systematic review and meta-analysis. Phytother Res. Mar 2019;33(3):473-481.
81. Walker AF et al. Bromelain reduces mild acute knee pain and improves well-being in a dose-dependent fashion in an open study of otherwise healthy adults. Phytomedicine. Dec 2002;9(8):681-6.
82. Jayachandran S et al. Efficacy of Bromelain along with Trypsin, Rutoside Trihydrate Enzymes and Diclofenac Sodium Combination Therapy for the treatment of TMJ Osteoarthritis - A Randomised Clinical Trial. Journal of clinical and diagnostic research : JCDR. Jun 2017;11(6):ZC09-ZC11.
83. Italiano G et al. Benefits of a Food Supplement Containing Boswellia serrata and Bromelain for Improving the Quality of Life in Patients with Osteoarthritis: A Pilot Study. Journal of alternative and complementary medicine (New York, NY). Feb 2020;26(2):123-129.
84. National Institute on Deafness and Other Communication Disorders. Taking Care of Your Voice. Updated 4/15/2021. https://www.nidcd.nih.gov/health/taking-care-your-voice
85. Centre ICE. The voice and its disorders in teachers. INSERM Collective Expert Reports. Institut national de la santé et de la recherche médicale. Copyright © 2000-, Institut national de la santé et de la recherche médicale (INSERM).
86. Baylor University. How to Prevent and Repair Vocal Damage. Updated 3/26/2020, https://onlinegrad.baylor.edu/resources/how-to-prevent-and-repair-vocal-damage-for-teachers-fitness-instructors-podcast-hosts-more/
87. Zambito P et al. THE EFFICACY OF EVAN® IN POST-OPERATIVE TREATMENT OF BENIGN CORDAL INJURIES: AN OBSERVATIONAL PILOT STUDY. Accessed 09/28/2021, http://www.frontieraorl.it/en/772-2/
88. Hancock RL et al. Vocal fold hemorrhage: when vocal fold blood vessels are not up to the task! Voice and Speech Review. 2014/01/02 2014;8(1):36-39.
89. Al-Ali M et al. The role of steroid injection for vocal folds lesions in professional voice users. J Otolaryngol Head Neck Surg. Jul 20 2020;49(1):50.
90. Insuan O et al. Anti-Inflammatory Effect of Pineapple Rhizome Bromelain through Downregulation of the NF-kappaB- and MAPKs-Signaling Pathways in Lipopolysaccharide (LPS)-Stimulated RAW264.7 Cells. Current issues in molecular biology. May 7 2021;43(1):93-106.
91. Paknezhad H et al. Vocal Fold Hemorrhage. Ear, Nose & Throat Journal. 2021/07/01 2019;100(6):NP276-NP277.
92. Shimizu S et al. Sesamin is a potent and specific inhibitor of delta 5 desaturase in polyunsaturated fatty acid biosynthesis. Lipids. 1991;26(7):512–516
93. Harikumar KB et al. Sesamin manifests chemopreventive effects through the suppression of NF-kappa B-regulated cell survival, proliferation, invasion, and angiogenic gene products. Mol. Cancer Res. 2010;8(5):751–761
94. Chavali SR et al. Decreased production of interleukin-1-beta, prostaglandin-E2 and thromboxane-B2, and elevated levels of interleukin-6 and -10 are associated with increased survival during endotoxic shock in mice consuming diets enriched with sesame seed oil supplemented with Quil-A saponin. Int. Arch. Allergy Immunol. 1997;114(2):153–160
95. Wu et al. Inhibition of 20-hydroxyeicosatetraenoic acid synthesis using specific plant lignans: in vitro and human studies. Hypertension. 2009;54(5):1151–1158
96. Miyawaki T et al. Antihypertensive effects of sesamin in humans. J. Nutr. Sci. Vitaminol. 2009;55(1):87–91
97. Srinivasan K. Cumin (Cuminum cyminum) and black cumin (Nigella sativa) seeds: traditional uses, chemical constituents, and nutraceutical effects. Food Quality and Safety. 2018;2(1):1-16.
98. Padhye S et al. From here to eternity - the secret of Pharaohs: Therapeutic potential of black cumin seeds and beyond. Cancer Ther. 2008;6(b):495-510.
99. Chung HY et al. Molecular inflammation: underpinnings of aging and age-related diseases. Ageing Res Rev. 2009;8(1):18-30.
100. Salem ML. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int Immunopharmacol. 2005;5(13-14):1749-1770.
101. Salem ML et al. Protective effect of black seed oil from Nigella sativa against murine cytomegalovirus infection. Int J Immunopharmacol. 2000;22(9):729-740.
102. Gheita TA et al. Effectiveness of Nigella sativa oil in the management of rheumatoid arthritis patients: a placebo controlled study. Phytotherapy research: PTR. 2012;26(8):1246-1248.
103. Nikakhlagh S et al. Herbal treatment of allergic rhinitis: the use of Nigella sativa. American journal of otolaryngology. 2011;32(5):402-407.
104. Al-Naqeep G et al. Antiatherogenic Potential of Nigella sativa Seeds and Oil in Diet-Induced Hypercholesterolemia in Rabbits. Evidence-based complementary and alternative medicine: eCAM. 2011;2011:213628.
105. Sourris KC et al. Ubiquinone (coenzyme Q10) prevents renal mitochondrial dysfunction in an experimental model of type 2 diabetes. Free Radic Biol Med. 2012 Feb 1;52(3):716-23.
106. Tao R et al. Pyrroloquinoline quinone preserves mitochondrial function and prevents oxidative injury in adult rat cardiac myocytes. Biochem Biophys Res Commun. 2007 Nov 16;363(2):257-62.
107. Rucker R et al. Potential physiological importance of pyrroloquinoline quinone. Altern Med Rev. 2009 Sep;14(3):268-77.
108. Xiong XH et al. Production and radioprotective effects of pyrroloquinoline quinone. Int J Mol Sci. 2011;12(12):8913-23.
109. Bauerly K et al. Altering pyrroloquinoline quinone nutritional status modulates mitochondrial, lipid, and energy metabolism in rats. PLoS One. 2011;6(7):e21779.
110. Donnino MW et al. Coenzyme Q10 levels are low and may be associated with the inflammatory cascade in septic shock. Crit Care. 2011 Aug 9;15(4):R189.
111. Sohet FM et al. Coenzyme Q10 supplementation lowers hepatic oxidative stress and inflammation associated with diet-induced obesity in mice. Biochem Pharmacol. 2009 Dec 1;78(11):1391-400.
112. Schmelzer C et al. Functions of coenzyme Q10 in inflammation and gene expression. Biofactors. 2008;32(1-4):179-83.
113. Stracke H et al. A benfotiamine-vitamin B combination in treatment of diabetic polyneuropathy. Exp Clin Endocrinol Diabetes. 1996;104(4):311-6.
114. Stracke H et al. Benfotiamine in diabetic polyneuropathy (BENDIP): results of a randomised, double blind, placebo-controlled clinical study. Exp Clin Endocrinol Diabetes. 2008 Nov;116(10):600-5.
115. Sanchez-Ramirez GM et al. Benfotiamine relieves inflammatory and neuropathic pain in rats. Eur J Pharmacol. 2006 Jan 13;530(1-2):48-53.
116. Shoeb M et al. Anti-inflammatory effects of benfotiamine are mediated through the regulation of the arachidonic acid pathway in macrophages. Free Radic Biol Med. 2012 Jan 1;52(1):182-90.
117. Vistoli G et al. Transforming dietary peptides in promising lead compounds: the case of bioavailable carnosine analogs. Amino Acids. 2012 Jul;43(1):111-26.
118. Fleisher-Berkovich S et al. Inhibitory effect of carnosine and N-acetyl carnosine on LPS-induced microglial oxidative stress and inflammation. Peptides. 2009 Jul;30(7):1306-12.
119. Tsai SJ. Antioxidative and Anti-Inflammatory Protection from Carnosine in the Striatum of MPTP-Treated Mice. J Agric Food Chem. 2010 Oct 6.
120. Boldyrev AA et al. [Carnosine: endogenous physiological corrector of antioxidative system activity]. Usp Fiziol Nauk. 2007 Jul-Sep;38(3):57-71.
121. Hipkiss AR. On the enigma of carnosine’s anti-ageing actions. Exp Gerontol. 2009 Apr;44(4):237-42.
122. Gualano B et al. Reduced muscle carnosine content in type 2, but not in type 1 diabetic patients. Amino Acids. 2011 Nov 27.
123. Pfister F et al. Oral carnosine supplementation prevents vascular damage in experimental diabetic retinopathy. Cell Physiol Biochem. 2011;28(1):125-36.
124. Gauhar R et al. Heat-processed Gynostemma pentaphyllum extract improves obesity in ob/ob mice by activating AMP-activated protein kinase. Biotechnology letters. 2012;34(9):1607-1616.
125. Winder WW et al. AMP-activated protein kinase, a metabolic master switch: possible roles in type 2 diabetes. Am J Physiol. 1999 Jul;277(1 Pt 1):E1-10.
126. Ruderman NB et al. AMPK, insulin resistance, and the metabolic syndrome. J Clin Invest. 2013 Jul 1;123(7):2764-72.
127. Park SH et al. Antiobesity effect of Gynostemma pentaphyllum extract (actiponin): a randomized, double-blind, placebo-controlled trial. Obesity (Silver Spring). 2014 Jan;22(1):63-71.
128. Towler MC, Hardie DG. AMP-activated protein kinase in metabolic control and insulin signaling. Circ Res. 2007 Feb 16;100(3):328-41.
129. Salminen A et al. AMP-activated protein kinase inhibits NF-kappaB signaling and inflammation: impact on healthspan and life span. J Mol Med (Berl). 2011 Jul;89(7):667-76.
130. Bijland S et al. Role of AMP-activated protein kinase in adipose tissue metabolism and inflammation. Clin Sci (Lond). 2013 Apr;124(8):491-507.
131. Xie Z et al. Chemical composition and anti-proliferative and anti-inflammatory effects of the leaf and whole-plant samples of diploid and tetraploid Gynostemma pentaphyllum (Thunb.) Makino. Food chemistry. 2012;132(1):125-133.
132. Bastard J-P et al. Recent advances in the relationship between obesity, inflammation, and insulin resistance. Eur. Cytokine Netw. 2006;17(1):4–12
133. Huyen VT et al. Antidiabetic effect of Gynostemma pentaphyllum tea in randomly assigned type 2 diabetic patients. Horm Metab Res. 2010 May;42(5):353-7.
134. Umeno A et al. Antioxidative and Antidiabetic Effects of Natural Polyphenols and Isoflavones. Molecules. May 30 2016;21(6).
135. Devi KP et al. Hesperidin: A promising anticancer agent from nature. Industrial Crops and Products. 2015;76:582-589.
136. Li C et al. Health-promoting effects of the citrus flavanone hesperidin. Critical reviews in food science and nutrition. Feb 11 2017;57(3):613-631.
137. Roohbakhsh A et al. Neuropharmacological properties and pharmacokinetics of the citrus flavonoids hesperidin and hesperetin--a mini-review. Life sciences. Sep 15 2014;113(1-2):1-6.
138. Jia S et al. Hypoglycemic and hypolipidemic effects of neohesperidin derived from Citrus aurantium L. in diabetic KK-A(y) mice. Food Funct. Mar 2015;6(3):878-886.
139. Rizza S et al. Citrus polyphenol hesperidin stimulates production of nitric oxide in endothelial cells while improving endothelial function and reducing inflammatory markers in patients with metabolic syndrome. The Journal of clinical endocrinology and metabolism. May 2011;96(5):E782-792.
140. Zhang J et al. Purification of naringin and neohesperidin from Huyou (Citrus changshanensis) fruit and their effects on glucose consumption in human HepG2 cells. Food chemistry. Dec 01 2012;135(3):1471-1478.
141. Homayouni F, Haidari F, Hedayati M, Zakerkish M, Ahmadi K. Hesperidin Supplementation Alleviates Oxidative DNA Damage and Lipid Peroxidation in Type 2 Diabetes: A Randomized Double-Blind Placebo-Controlled Clinical Trial. Phytotherapy research. Aug 14 2017.
142. Sun Q et al. Urinary Excretion of Select Dietary Polyphenol Metabolites Is Associated with a Lower Risk of Type 2 Diabetes in Proximate but Not Remote Follow-Up in a Prospective Investigation in 2 Cohorts of US Women. The Journal of nutrition. Jun 2015;145(6):1280-1288.
143. Salden BN et al. Randomized clinical trial on the efficacy of hesperidin 2S on validated cardiovascular biomarkers in healthy overweight individuals. The American journal of clinical nutrition. Dec 2016;104(6):1523-1533.
144. Haidari F et al. Hesperidin supplementation modulates inflammatory responses following myocardial infarction. Journal of the American College of Nutrition. 2015;34(3):205-211.
参考来源:
美国国立卫生研究院
http://www.nih.gov
美国国立公众健康网
www.medlineplus.gov
英国今日医学新闻网
www.medicalnewstoday.com
免责声明和安全信息
- 本信息(包括任何附带资料)不是为了取代医生或有关合格从业人士的建议或忠告。
- 任何人如果想要对本文涉及的药物、饮食、运动或其他生活方式的使用、或改变调整,以预防或治疗某一特定健康状况或疾病,应首先咨询医生或有关合格从业人士,并获得他/她们的许可。妊娠和哺乳妇女在使用本网站任何内容前,尤其应征求医生的意见。
- 除非另有说明,本网站所述内容仅适用于成人。
- 本网站所推荐的任何产品,消费者应该以实际的产品标签内容为准,尤其应关注重要的安全信息以及产品最新信息,包括剂量、使用方法和禁忌症等。
- 由于循证医学研究、文献及有关产品处于不断的变化中,本网站工作人员将尽力更新。
- 本网站不能保证所载文章内容、综合干预方案以及相关成分或产品述及的健康益处,也不承担任何责任。